

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
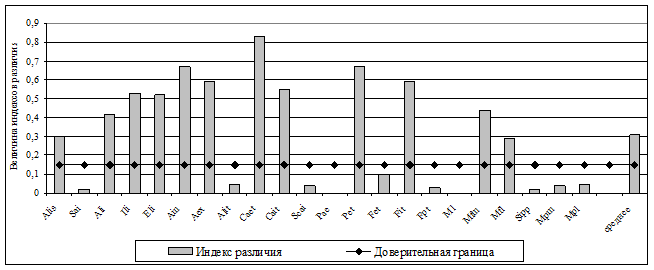
Более подробно исследована динамика фенетического разнообразия рыжих полевок в гг. в коренных неморальных ельниках. В этот период укладывается несколько популяционных волн: два полных цикла с типичным S-образным ростом ( гг., гг.) и начало третьего цикла (см. рис 9). Полученные результаты указывают на достаточно высокую степень фенетических отличий популяционных группировок в различные годы, как по отдельным признакам, так и по их комплексу (рис. 12).
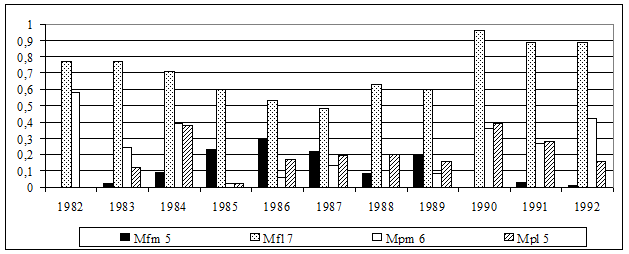
Рис. 12. Динамика частоты встречаемости некоторых фенов в группе вариабельных краниологических признаков
Обнаружено, что наиболее существенные перестройки феногенетического состава происходили на последней фазе 4-5 летних популяционных циклов (1985, 1990 гг.) перед снижением численности (рис. 13). Причем, степень фенетического сходства выборок последнего года популяционных циклов была несколько выше с начальной фазой следующего цикла, чем с предшествующим годом своего цикла. В рамках одного цикла прослеживаются закономерные изменения частоты встречаемости отдельных фенов в зависимости от уровня численности. Отмечена определенная периодичность формирования фенетической структуры популяции рыжей полевки в ходе многолетней динамики численности (см. рис. 12), что может свидетельствовать об устойчивости феногенетического разнообразия вида в коренных лесах.
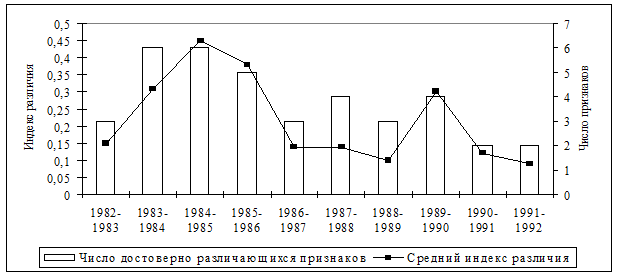
Рис. 13. Динамика степени различий фенетической структуры популяций рыжей полевки
Для оценки влияния на динамику фенетической структуры процесса воспроизводства и выживаемости были сформированы серии черепов перезимовавших животных и неполовозрелых сеголеток из осенних отловов (октябрь), уходящих на зимовку за период гг. Обнаружено, что специфика воспроизводства может определять довольно быстрые и значимые изменения феногенетической структуры в ходе одного репродуктивного сезона. Наиболее существенные перестройки зарегистрированы в сезон с активным участием в размножении сеголеток ранних сроков рождения. При этом в определенной степени в репродукции выражен возрастной кросс, а осенние выборки неполовозрелых особей состоят из двух поколений. Выявлены изменения фенетической структуры в зависимости от условий перезимовки популяции. При сравнении осенних и весенних выборок для ряда признаков отмечены вариации, устойчиво из года в год демонстрирующие увеличение доли своего присутствия от осени к весне. Характер изменения частоты встречаемости данных фенов не зависел ни от специфики климатических условий, ни от плотности популяции. Это косвенно может свидетельствовать о хорошей выживаемости особей-носителей данных фенов, которые можно считать своеобразными маркерами животных с высокой неизбирательной жизнеспособностью. В то же самое время были встречены фены, частота которых в популяции от осени к весне следующего года менялась неодинаковым, но вполне закономерным образом, демонстрируя определенную связь со степенью благоприятности условий перезимовки. В свою очередь итоговая смертность популяции, безусловно, зависит от конкретного соотношения феногенотипов, имеющих неодинаковую выживаемость при различных сочетаниях погодных и других факторов. Указанные обстоятельства, несомненно, должны учитываться при анализе и прогнозировании динамики популяций.
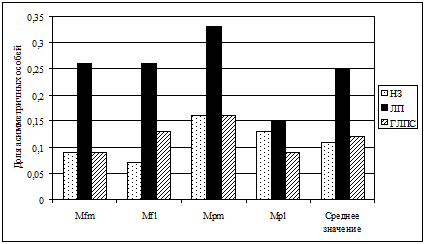
В настоящее время практически не изучены аспекты феногенетики переносчиков зоонозных заболеваний на внутри - и межпопуляционном уровнях. В работе предпринимается попытка фенетического тестирования особей рыжей полевки по степени их восприимчивости и инфекционной чувствительности к основным возбудителям природно-очаговых болезней региона и оценивается влияние эпизоотий на фенетическое разнообразие популяций вида. Обнаружено, что фенетическая структура группировок особей, зараженных лептоспирозом, достоверно отличалась от таковой неинфицированных особей. Группировка особей, инфицированных ГЛПС, имела меньший уровень отличий от незараженных животных и величина среднего значения фенетических дистанций не достигала порогового уровня значимости. Отмечено снижение доли редких фенов в инфицированных группах особей. В целом, гораздо более существенно воздействие лептоспирозов, поскольку вмешивается фактор патогенеза при заражении особей, который снижает стабильность их индивидуального развития (рис. 14) и может увеличивать смертность конкретных феногенотипов в популяции. Полученные результаты дают основание предполагать, что феногенетическая структура популяций основного носителя инфекций может оказывать существенное воздействие на общий ход течения эпизоотии. Это, по-видимому, связано с различной инфекционной чувствительностью определенных феногенотипов особей.
| Рис. 14. Доля асимметричных особей в инфицированных и неинфицированных группировках рыжей полевки по наиболее полиморфным признакам; НЗ – незараженные особи, ЛП – инфицированные лептоспирозом; ГЛПС – инфицированные геморрагической лихорадкой. |
Для мышевидных грызунов характерна сложная функциональная структурированность популяций. В данном разделе рассматриваются фенетические особенности оседлых и расселяющихся рыжих полевок, а также особей с различными типами онтогенеза. Роль указанных структурно-функциональных группировок грызунов в динамике ведущих популяционных параметров, безусловно, очень велика.
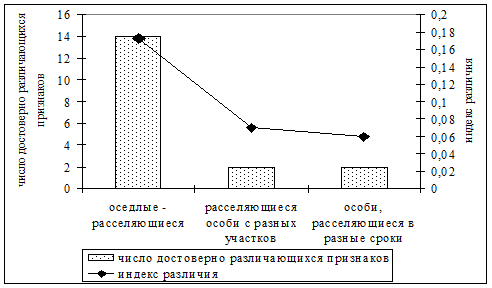
Процесс расселения во многом определяет пространственную структуру и жизнеспособность популяций. Он чрезвычайно сложен для исследований, поэтому существует много нерешенных проблем по разным его аспектам (Myers, Krebs, 1971; Hilborn, 1975; Fairbairn, 1978; Коли, 1979; Staffard, Staout, 1983; Gliwicz, 1986; Rease et all, 1989; Лукьянов, Лукьянова, 1996; Лукьянов, 1997 и др.). Генетическая обусловленность расселения является одним из наименее изученных вопросов. Нами установлено, что в континуальной популяции рыжей полевки группировки оседлых и расселяющихся особей фенетически разнородны (рис. 15). В то же самое время различия фенетической структуры расселяющихся особей, отловленных на разных участках и в разные периоды расселения, выражены несущественно и носят недостоверный характер. Это может косвенно свидетельствовать об определенной генетической предрасположенности особей к расселению. Обнаружено, что довольно активно расселяются массовые фенотипы. Однако для некоторых немногочисленных у оседлых особей вариаций отмечено существенное увеличение частоты их встречаемости в группировках расселяющихся особей. Вероятно, именно эти признаки и фены могут быть рекомендованы в качестве маркировочных при изучении процессов дисперсии в популяциях рыжей полевки.
| Рис. 15. Число достоверно различающихся признаков (из 24) и величина индекса различия между сравниваемыми группировками оседлых (n=166) и расселяющихся (n=160) особей |
Темпы роста численности популяций грызунов во многом определяются степенью участия в репродукции сеголеток. Этот показатель очень изменчив, что связано с наличием двух альтернативных путей онтогенеза: однофазный рост и двухфазный рост (Шварц, 1980; Большаков и др., 1991; Оленев, 1991 и др.). В результате этого в популяции имеются разные функционально-физиологические группы особей (ФФГ). В изучаемой нами популяции рыжей полевки доля особей с однофазным ростом, размножающихся в год своего рождения (3 ФФГ), варьировала в широких пределах% - для самцов, 1 - 55% – для самок). У данной группы, прежде всего у самцов, обнаружен более высокий уровень флуктуирующей асимметрии качественных и количественных признаков. Это свидетельствует о снижении стабильности развития особей с однофазным ростом, что в определенной степени может уменьшать их жизнеспособность и сокращать продолжительность жизни. У перезимовавших животных (1ФФГ, бывшая 2ФФГ) доля асимметричных особей становится еще меньше, чем у их предшественников. Так, в случае с комплексным признаком непарная петля параконоида в суммарной группировке ( гг.) перезимовавших особей, отмечено только 7,6% асимметричных особей. Тогда как во 2ФФГ суммарная доля асимметричных особей составляла 15,8%, а у особей с однофазным ростом (3ФФГ) – 19,7%. Эти результаты дополнительно подтверждают предположение о взаимосвязи стабильности онтогенеза и общей продолжительности жизни особей.
Популяционная динамика красной полевки в коренных бореальных ельниках. Красная полевка, являясь восточно-палеарктическим видом, в лесах южной тайги предпочитает бореальные ельники, где образует постоянные полуизолированные группировки. Изучали динамику численности красной полевки на двух крупных участках олиготрофных бореальных ельников, один из которых в конце 80-х - начале 90-х гг. подвергся частичному разрушению через групповое усыхание ели и последующее ее вываливание. Модельные площади находятся на расстоянии приблизительно 1 км друг от друга и изолированы лесными местообитаниями, в целом не свойственными красной полевке (неморальные ельники и их производные). Многолетние изменения численности вида на сравниваемых участках в целом аналогичны (r=0,75) и отражают нестационарные процессы с положительными трендами и периодическими составляющими (рис. 16).
|
Рис. 16. Многолетняя динамика и тренды численности красной полевки в коренных бореальных ельниках заповедника; достоверность аппроксимации для участка 1 R^2=0,57; для участка 2 R^2=0,48. |
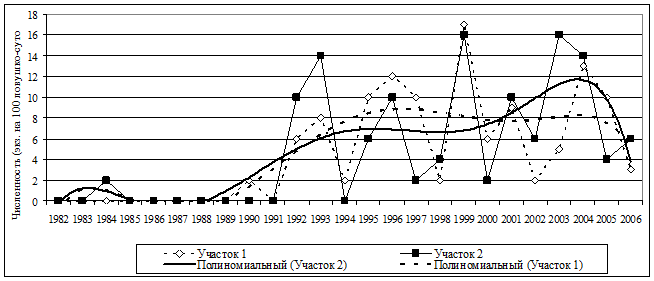
На долю трендов приходится приблизительно 50-57% варьирования данных. Достаточно хорошо исходные временные ряды аппроксимируются полиномиальными кривыми шестой степени. Выделяются два периода ( г. г.; г. г.) с разным уровнем и характером динамики численности. Средние значения летней численности сравниваемых периодов различаются в 36 раз. Тестирование первой части ряда показало, что, несмотря на относительную стационарность процесса, имеется слабо выраженная периодическая составляющая, связанная с внутригодовыми колебаниями, а регулярные многолетние колебания отсутствуют. Результаты тестирования второй части ряда свидетельствуют о незначительном положительном тренде и регулярных составляющих в динамике численности продолжительностью 3-4 года. При построении модели с учетом выявленного 4-летнего цикла получены достаточно высокие коэффициенты детерминации, которые указывают, что вклад указанной составляющей в общее варьирование данной части ряда составляет 16-42%. Переход между двумя различными режимами популяционной динамики произошел достаточно резко. Зона перехода из одного режима в другой вполне может рассматриваться как зона нарушения относительной стационарности процесса. Причем, в отличие от рыжей полевки, для которой в исследованный промежуток времени на фоне увеличения численности наблюдалось явное упрощение популяционных циклов, у красной полевки происходило усложнение характера цикличности популяций.
Исследованы особенности поддержания феногенетического разнообразия реликтового вида в естественных южно-таежных лесах. Описана фенетическая структура отдельных пространственных группировок, определены фенетические дистанции между ними, даются оценки величины и структуры фенетического разнообразия. Популяционная система красной полевки в условиях коренных лесов заповедника полиморфна (для 33 признаков черепа выявлено 118 неметрических вариаций). Это свидетельствует о достаточно высокой жизнеспособности и микроэволюционной активности данного реликтового вида. При изоляции в пределах 1 км без явных эколого-географических барьеров суммарная степень фенетической разнородности популяционных группировок относительно невелика, что косвенно указывает на существование регулярного генетического обмена между группировками, которые изолированы сходными дистанциями. Однако структура феногенетического разнообразия отдельных популяционных группировок может иметь определенные отличия. Так, в ненарушенном ельнике существенный вклад в суммарный показатель разнообразия вносят редкие морфы, встречаемость которых в ельнике с частичными нарушениями несколько ниже. Наоборот, показатели внутрииндивидуального разнообразия выше в ельнике с нарушениями. Это свидетельствует о некотором снижении стабильности онтогенеза особей красной полевки и совершенства его регуляторных процессов в дестабилизированных лесных экосистемах, по сравнению с климаксными бореальными ельниками.
Популяционная динамика полевки-экономки на ранних стадиях демутации ельников. Полевка-экономка - обычный, но спорадически распространенный вид, формирующий сложную, чрезвычайно динамичную в пространстве и во времени популяционную систему, которая состоит из достаточно изолированных и эфемерных группировок, приуроченных к самым ранним стадиям демутаций ельников. Исследовано несколько колонизационных циклов (заселение, нарастание численности, популяционный расцвет и крах) полевки-экономки на крупных участках массовых вывалов и вырубках. Обнаружено, что циклы реализуются в короткие сроки (5-7 лет). Кривые динамики численности полевки-экономки на различных ветровальных участках в целом имели достаточно сходный характер (рис. 17).
| Рис. 17. Динамика численности полевки-экономки в ходе колонизационных циклов на массовых вывалах и сплошной вырубке |
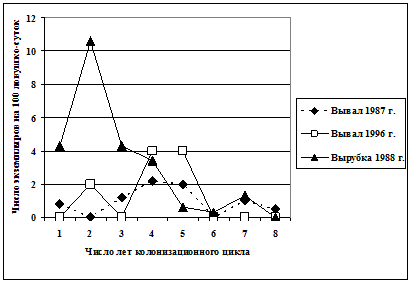
Существенно более высокими были показатели численности вида на вырубках. Колонизационные циклы хорошо аппроксимируются полиномиальными трендами шестой степени. Отмечен общий характер сезонных изменений численности в ходе колонизационных циклов: максимальные значения регистрируются в начале-середине лета. Обнаружена асинхронность движения численности группировок вида на вывалах и вырубках, даже появившихся в одно и то же время (r=-0,26). Это может означать, что популяционная динамика отдельных группировок достаточно независима.
Подробно исследованы пространственно-временные особенности фенетической структуры двух популяционных группировок, населяющих ранние стадии вырубок, основные характеристики которых (исходное сообщество, время и характер рубки, размер вырубленного участка и характер нарушенности) практически идентичны. Всего для 22 признаков выявлен 71 фен. Сравниваемые группировки фенетически очень разнородны (рис. 18), что дает основание предполагать о существовании определенных генетических различий между ними и первостепенной роли основателей при формировании феногенетического разнообразия популяций пионерных видов.
|
Рис. 18. Фенетические дистанции между исследованными популяционными группировками полевки-экономки |
Установлено, что фенетическая структура короткоживущей популяции полевки-экономки чрезвычайно динамична во времени. Степень реализации локального фенофонда существенно возрастала на фазах колонизационного расцвета, продолжая оставаться достаточно высокой и в начале периода сокращения численности популяции.
Разные фазы популяционного цикла достоверно отличались по частотам фенов многих признаков. Суммарная степень различий фенетической структуры увеличивалась от момента заселения до завершения основной части колонизационного цикла (рис. 19).
| Рис. 19. Динамика степени различий фенетической структуры популяций полевки-экономки в ходе колонизационного цикла |
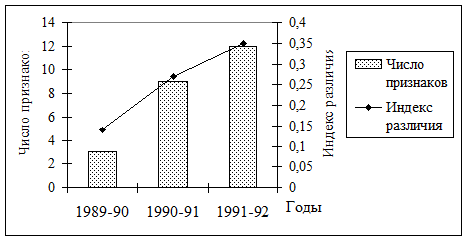
Это может свидетельствовать об усилении процессов дифференциации в популяционных системах полевки-экономки в ходе колонизационного цикла, что, безусловно, приводит к увеличению феногенетического разнообразия вида на данной территории. Общая численность вида, характер ее изменений и феногенетическое разнообразие определяются, в первую очередь, динамикой и масштабами ветровальных процессов и сплошных рубок. Несмотря на сходство колонизационных циклов, популяционная динамика отдельных эфемерных группировок полевки-экономки достаточно независима, а фенетическое разнообразие и структура субпопуляций специфичны.
Выводы
1. В фауне мелких млекопитающих естественных лесов центра Каспийско-Балтийского водораздела преобладают зональные виды. В стабильном и жизнеспособном состоянии находятся популяции реликтовых видов. Лесостепные, синантропные, интродуцированные виды немногочисленны или совсем отсутствуют. Это может указывать на экологическую полночленность природных сообществ, их способность противостоять агрессивному воздействию и развитию чужеродной биоты.
2. В коренных ельниках разного генезиса центра Каспийско-Балтийского водораздела в последние 15 лет существенно возросли альфа-разнообразие и суммарное обилие гильдий мелких млекопитающих, а также их зональная контрастность и «гетероморфность», что свидетельствует об увеличении бета-разнообразия сообществ в лесном массиве в целом.
3. Сообщества мышевидных грызунов коренных ельников сходным образом реагируют на воздействие массовых ветровалов и сплошных рубок, формируя на ранних стадиях демутаций высокое видовое и трофическое разнообразие. В ходе послерубочной сукцессии в первые годы наблюдается быстрая смена доминирования видов, а коренная структура гильдий грызунов восстанавливается в возрасте 60-80 лет, когда сообщества ельников вступают в устойчивую фазу развития.
4. Динамика численности ценозообразующих видов полевок (рыжей и красной) коренных ельников в гг. представляла собой нестационарный процесс с существенным вкладом в общее варьирование положительного тренда и цикличности. Произошли изменения амплитуды колебаний, продолжительности циклов, типов кривых популяционного роста. В различных типах естественных лесных экосистем численность видов изменялась достаточно синхронно. На ранних стадиях вырубок, по сравнению с коренными лесами, обилие рыжей полевки снижается, изменяется характер многолетней и сезонной динамики численности.
5. Фенетическая структура популяций фоновых видов полевок коренных ельников изменяется в зависимости от фаз циклов и уровней формируемой численности, условий перезимовки и особенностей воспроизводства. Наиболее существенные перестройки феногенетического состава рыжих полевок происходят на последней фазе циклов перед снижением численности. В рамках отдельных циклов прослеживаются закономерные изменения частот встречаемости определенных фенов.
6. В крупном массиве естественных лесов фенетические различия между пространственными группировками континуальной популяции рыжей полевки в диапазоне удаленностей до 4 км сравнительно невелики, а связь между величиной фенетической дистанции и изоляцией расстоянием отсутствует. Фенетические различия возрастают в годы пониженной численности вида. В подразделенных популяциях рыжей полевки увеличивается общее фенотипическое разнообразие. Это происходит, в первую очередь, за счет увеличения уровня случайных изменений, связанных с последствиями инсуляризации.
7. На участках массовых ветровалов и сплошных вырубок сходным образом изменяется фенетическая структура популяций рыжей полевки. Как правило, сокращается общее популяционное разнообразие и доля редких морф. Возрастают показатели флуктуирующей асимметрии, и уменьшается положительная скоррелированность развития признаков. Это свидетельствует о снижении стабильности онтогенеза и совершенства его регуляторных процессов в условиях экологически дестабилизированных сред ранних стадий демутации и указывает на возможность использования в биомониторинге таких универсальных явлений как изменчивость и развитие организмов.
8. Внутрипопуляционные функциональные группы особей рыжей полевки фенотипически разнородны. Достоверно различается фенетическая структура оседлых и расселяющихся рыжих полевок. Особи с разными типами онтогенеза имеют неодинаковые уровни флуктуирующей асимметрии количественных и качественных признаков. Более высокие показатели асимметрии животных с однофазным ростом указывает на снижение стабильности их индивидуального развития.
9. В центре Каспийско-Балтийского водораздела функционируют активные лесные очаги лептоспирозов и ГЛПС, которые влияют на динамику фенетической структуры и разнообразия популяций основных носителей инфекций. В инфицированных группах снижается доля редких фенов. Более существенно воздействие лептоспирозов, поскольку вмешивается фактор патогенеза при заражении особей.
10. Активная сукцессионная динамика лесов приводит к усложнению популяционных систем пионерных видов грызунов, которые формируют на ранних стадиях демутаций довольно многочисленные эфемерные группировки. Цикл колонизации полевки-экономки реализуется в течение 5-7 лет. Популяционная динамика отдельных группировок достаточно независима, а фенетическая структура специфична. В ходе колонизационных циклов усиливаются процессы дифференциации, что приводит к увеличению феногенетического разнообразия вида.
11. Популяции и сообщества мелких млекопитающих демонстрируют закономерные отклики на изменения лесных экосистем, возникающие в ходе антропогенных воздействий и естественных процессов различного характера и масштаба, что подтверждает их биоиндикационную значимость для комплексного экологического мониторинга.
СПИСОК ОСНОВНЫХ ОПУБЛИКОВАННЫХ РАБОТ ПО ТЕМЕ ДИССЕРТАЦИИ
Монографии
1. Истомин фенетика рыжей полевки (на примере южной тайги). Монография. Псков, 20с. (12,25 п. л.).
2. Истомин млекопитающие в региональном экологическом мониторинге (на примере Каспийско-Балтийского водораздела). Монография. Псков, 20с. (17,4 п. л.).
3. Методические рекомендации по ведению мониторинга на особо охраняемых природных территориях (на примере Центрально-Лесного государственного природного биосферного заповедника). Коллективная монография. М., 20с. (14,5 п. л., авторский вклад – 30%).
Научные статьи, материалы конференций, совещаний
4. Истомин видов и характер распространения мелких млекопитающих на островах Кандалакшского залива в Белом море // Зоологический журнал. Т. 63, №С. . (1,0 п. л.).
5. Истомин разнообразие континуальной и дискретной популяций на примере рыжей полевки в условиях южной тайги // Журнал общей биологии. Т. 55, № 4С. 471—488. (1,0 п. л.).
6. Истомин межвидовой конкуренции в формировании островных сообществ мелких млекопитающих // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 99, вып.С. 17-26. (1,0 п. л.).
7. , , Ананьина полевка (Clethrionomys glareolus) — основной носитель лептоспироза в лесном природном очаге // Зоологический журнал. Т. 72. Вып.С. 113—122. (1,0 п. л., - авторский вклад – 30%).
8. Истомин накопления радионуклидов в тканях рыжих полевок из природных популяций фоновой территории центра Русской равнины // Вестник РУДН. Серия «Экология и безопасность жизнедеятельности». 2009. № 1. С. 75-80. (0,4 п. л.).
9. Истомин ветровалов на динамику сообществ мелких млекопитающих в естественных лесах южной тайги // Вестник Московского государственного университета леса – Лесной вестник. 2009. №1 (64). С. 196-202. (0,6 п. л.).
10. Истомин экологической дестабилизации среды сплошных вырубок на онтогенез лесных видов мелких млекопитающих // Вестник РУДН. Серия «Экология и безопасность жизнедеятельности». 2009. № 2. С. 9-16. (0,4 п. л.).
11. Истомин распространения и биологии подземной полевки на северо-востоке ареала // Вестн. зоол., Киев, 1990, № 3. С. 61-63. (0,5 п. л.).
12. Истомин полидонтии у лесной рыжей полевки (Clethrionomys glareolus) из двух пунктов ареала // Вестн. зоол., 1994. Киев, № 1. С. 83-86. (0,5 п. л.).
13. Истомин инвентаризационных исследований фауны мелких млекопитающих в заповедниках // Теоретические основы заповедного дела: Тез. докл. Всесоюз. совещ., Львов, 1985. С. 108—111. (0,2 п. л.).
14. Истомин экология и теория заповедного дела // Там же. С. 105-108. (0,2 п. л.)
15. , Мелкие млекопитающие как объекты биологического мониторинга лесных экосистем // Мониторинг лесных экосистем: Тез. докл. Всесоюз. научн. конф., Каунас, 1986. С. 95—96. (0,1 п. л.).
16. Истомин морфологической изменчивости рыжей европейской полевки в островных популяциях // 4 съезд ВТО. Москва, 1986. Т. 2, М., 1986. С. 51-52. (0,1 п. л.).
17. Истомин размеров тела и полового диморфизма популяций рыжей европейской полевки в островных условиях // Организмы, популяции и сообщества в экстремальных условиях. Всесоюзн. сов., Москва, 1986, М. С. 59-61. (0,1 п. л.).
18. Истомин сплошных рубок на популяционную динамику европейской рыжей полевки в условиях южной тайги // Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных: Всесоюз. сов., Москва, 1987. Ч. 2. М. С.18—20. (0,1 п. л.).
19. Истомин мелких млекопитающих в ходе послерубочной сукцессии неморальных ельников // Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных: Всесоюз. сов., Москва, 1987. Ч. 2. М. С. 63—64. (0,1 п. л.).
20. Истомин мелких млекопитающих южной тайги в условиях антропогенной трансформации ландшафтов // Животный мир лесной зоны европейской части СССР. Калинин, 1988. С. 37—45. (0,6 п. л.).
21. Истомин полевка - перспективный объект для разработки региональной программы в заповедниках лесной зоны Европейской части СССР // Научные исследования в заповедниках и принципы разработки региональных программ для заповедников лесной зоны Европейской части СССР. Тез. докл. Всесоюз. науч. конф. Рахов, 1990. С. 50—53. (0,2 п. л.).
22. Истомин инсуляризации и поддержание жизнеспособных популяций животных в резерватах // Заповедники СССР - их настоящее и будущее. Всесоюзн. конф., Ч. 3, Новгород, 1990. С. 240-243. (0,2 п. л.).
23. , Кораблев и методы охраны генофонда популяций млекопитающих и их реализация в ЦЛБЗ // Заповедники СССР - их настоящее и будущее. Всесоюзн. конф., Ч. 3, Новгород, 1990. С. 257-259. (0,2 п. л., авторский вклад – 50%).
24. Истомин индивидуального развития и возможность использования показателя флуктуирующей асимметрии при оценке устойчивости экосистем // Проблемы устойчивости экосистем: Тез докл. Всесоюз. школы. Харьков, 1990. С. 82—84. (0,2 п. л.).
25. , Алексеева фенов жевательной поверхности М1 европейской рыжей полевки на разных фазах динамики численности // Y съезд Всесоюз. Териолог. об-ва, Москва. Тез. докл. Т. 2, М., 1990. С. 156-157. (0,1 п. л. – авторский вклад - 80%).
26. Истомин экосистем южной тайги // Человек и биосфера: информационный бюллетень MAB. № 2. М., 1991. С. 17—20. (0,3 п. л.)
27. В, Вагин болота как ландшафтно-экологические барьеры: роль "болотных изолятов" в микроэволюционных процессах // Болота охраняемых территорий: проблемы охраны и мониторинга. 11 всесоюзн. полевой семинар-экскурсия по болотоведению. Л., 1991. С. 117-120. (0,3 п. л., авторский вклад 80%).
28. , , Шлыгина очаг туляремии лесного типа в Тверской области // Актуальные проблемы профилактики туляремии. Всесоюзн. конф., 15-17 окт. 1991 г., Симферополь, М., 1991. С.48-49. (0,1 п. л., авторский вклад – 20%).
29. Истомин нарушений на видовое разнообразие мышевидных грызунов в южной тайге // Видовое разнообразие млекопитающих в трансформированных экосистемах. - Киев: Ин-т зоологии Украины (Препринт 92.3), 1992. С. 29—40. (0,7 п. л.).
30. Истомин мышевидных грызунов в ходе вторичной антропогенной сукцессии ельников южной тайги - количественное сравнение // Фауна и экология животных лесной зоны. Тверь, 1992. С. 99—108.(0,6 п. л.).
31. Истомин разнообразие и микроэволюционное состояние популяций в различных системах резерватов // Некоторые проблемы изучения разнообразия млекопитающих. Киев: Ин-т зоологии АН Украины (Препринт 92.4), 1992. С. 26-35. (0, 6 п. л.).
32. Истомин населенных пунктов малоосвоенной территории водораздела Волги и Западной Двины // Синантропные грызуны. Матер. 2 совещ., Иваново, 1993. Характер поведения. Особенности обитания в постройках человека. Методы учетов. Пути ограничения численности. М., 1994. С. 124—129. (0,5 п. л.).
33. Истомин трансформация ландшафтов Волжско-Двинского водораздела и состояние природных комплексов Центрально-Лесного биосферного заповедника // Социальные и экономические аспекты заповедного дела. Междунар. совещ. С-Пб., 1994.С. 112—115. (0,2 п. л.).
34. Истомин и распространение лесной мыши в Верхневолжье // Фауна и экология животных Верхневолжья. Тверь, 1994. С. 88-94. (0,6 п. л.).
35. , , Блохи мелких млекопитающих Центрально-Лесного биосферного заповедника // Фауна и экология животных Верхневолжья. Тверь, 1994. С. 27-32. (0,6 п. л., авторский вклад – 50%).
36. The effect of disturbance on the small mammals communities and population diversity in the Southern taiga in European Russia // Convercing biological diversity in temperate forest ecosystems - towards sustainable management: Conference, Canberra-Australia, December 1994. (0,2 п. л.)
37. Истомин млекопитающие как объекты комплексной оценки биологического разнообразия лесных экосистем // Биологическое разнообразие лесных экосистем: Тез. докл. Всерос. Совещ. М., 1995. С. 144—146. (0,2 п. л.).
38. Истомин стадии "окон" в поддержании разнообразия сообществ мышевидных грызунов в коренных еловых лесах южной тайги // Биологическое разнообразие лесных экосистем: Тез. докл. Всерос. Совещ. М., 1995. С. 146—148.
39. Истомин Центрально-Лесного биосферного заповедника // Флора и фауна заповедников России. Позвоночные животные Центрально-Лесного заповедника. Вып. 59. М., 1995. С. 33—42. (0,6 п. л.).
40. Истомин оценка биологического разнообразия охраняемых природных территорий лесной зоны // Проблемы сохранения биоразнообразия Псковской области. СПб., Изд-во СПб. ун-та, 1998. С. 7—16. (0,6 п. л.).
41. «Бореализация» сообществ мелких млекопитающих в коренных южно-таежных лесах центра Русской равнины // Экология таежных лесов. Тез. докл. Межд. конф. Сыктывкар, 1998. С. 141—142. (0,1 п. л.).
42. Истомин колонизации пионерных видов и их роль в формировании генетического разнообразия // Циклы природы и общества. Матер. 7 междунар. конф. Ставрополь, 1999. С. 228—231. (0, 2 п. л.).
43. Истомин и опыт использования мелких млекопитающих в экологическом мониторинге Центрально-Лесного биосферного заповедника (ЦЛБЗ) // Экологический мониторинг лесных экосистем. Матер. Всерос. Совещ. Петрозаводск, 1999. С.14. (0,1 п. л.).
44. Истомин лептоспирозов в естественных и антропогенных ландшафтах Центрального Нечерноземья России // Актуальные вопросы биоразнообразия животных в антропогенном ландшафте. Тезисы докл. научно-практ. конф. Киев, изд-во УА МБН, 1999. С. 57-61. (0,2 п. л.).
45. Истомин и динамика численности полевки-экономки и рыжей полевки на ранних стадиях зарастания сплошных вырубок южной тайги // Актуальные вопросы биоразнообразия животных в антропогенном ландшафте. Тезисы докл. научно-практ. конф. Киев, изд-во УА МБН, 1999. С. 62-65. (0,2 п. л.).
46. Истомин и миграция загрязняющих веществ в лесных экосистемах Центрально-Лесного биосферного заповедника. Экологический мониторинг лесных экосистем. Материалы Всероссийского совещания. Петрозаводск, 1999. С. 43. (0,1 п. л).
47. Истомин лемминг - реликт южной тайги центра Русской равнины // Доклады третьих международных чтений памяти профессора , Вып.3. Смоленск, 2000. С. 237—239. (0,2 п. л.).
48. Истомин териокомплексов и оценка степени антропогенной трансформации территорий // Экология и рациональное природопользование на рубеже веков: итоги и перспективы. Тез. докл. межд. конф. Т 2. Томск, 2000. С.52—54. (0,2 п. л.)
49. Истомин анализ морфологической изменчивости популяций в целях биомониторинга охраняемых территорий (на примере грызунов) // Исследования на охраняемых природных территориях Северо-запада России. Матер. регион. науч. конф., посвящ. 10-летию Валдайского Национального парка. Великий Новгород, 2000. С. 306—307. (0,1 п. л.).
50. Истомин мышь в естественных и антропогенных ландшафтах Каспийско-Балтийского водораздела // Социальные и экологические проблемы Балтийского региона. Матер. обществ.-науч. конф. Псков, 2000. С.157—160. (0,2 п. л.).
51. Истомин охраняемых природных территорий и мониторинг последствий их инсуляризации // Особо охраняемые природные территории. 11 международная конференция. СПб., 2000. С.44-45. (0,1 п. л.).
52. , Мониторинг биологического разнообразия на охраняемых территориях // Теоретические основы биоразнообразия. Материалы семинара, СПбГУ, СПб, 2000. С. 41-42. (0,1 п. л.).
53. Истомин модели эпизоотий лептоспирозов в лесных очагах на территории Волжско-Двинского водораздела // Северо-Запад России: взаимодействие общества и природы. Материалы конференции с международным участием. Ч. 1. Псков, 2001. С.190-194. (0,2 п. л.).
54. Истомин -эпидемическое значение лесных ландшафтов на территории Каспийско-Балтийского водораздела // Социальные и экологические проблемы Балтийского региона. Материалы конференции с международным участием. Ч. 2. Псков, 2001. С. 265 – 276. (0,8 п. л.)
55. Истомин онтогенеза лесных видов в естественных и нарушенных экосистемах южной тайги // Проблемы изучения и охраны биоразнообразия и природных ландшафтов Европы. Материалы Международного симпозиума. Пенза, 2001. С. 176-179. (0,1 п. л.).
56. , , и др. К изучению реакции биоты Центрально-лесного заповедника на изменения климата // Влияние изменения климата на экосистемы. М.: Русский университет, 2001. С. 87-100. (3,0 п. л., авторский вклад – 30%).
57. Истомин компоненты и тренды в динамике численности популяций рыжей полевки в коренных бореальных ельниках южной тайги // Циклы. Материалы IY междунар. конф. Ставрополь, 2002. С. 82-84. (0,2 п. л.).
58. Истомин лесных ландшафтов сплошными рубками и формирование очагов лептоспироза // Природные и культурные ландшафты: проблемы экологии и устойчивого развития. Материалы конф. с междунар. участием. Статьи и тезисы. Ч. 1. Псков, 2002. С. 137-140. (0,2 п. л.).
59. Истомин проблемы создания экологических сетей (Econet) // РИО+10: Охрана окружающей среды, природопользование, образование. Материалы Псковской областной экологической конференции. Выпуск 7. Великие Луки, 2002. С. 129-134. (0,3 п. л.).
60. Истомин естественных местообитаний и функционирование охраняемых природных территорий (ОПТ) // РИО+10: Охрана окружающей среды, природопользование, образование. Материалы Псковской обл. экол. конф. Вып. 7. Великие Луки, 2002. С. 135-140. (0,3 п. л.).
61. В, Истомин – Mammalia // Красная книга Тверской области. Тверь: Твери», «Издательство АНТЭК», 2002. С. 153-161. (0,5 п. л. – авторский вклад 30%).
62. Истомин индивидуального развития особей с различными типами онтогенеза // Научные чтения памяти профессора , вып. 4. Смоленск: Изд-во СГПУ, 2004. С. 381-386. (0,4 п. л.).
63. , Истомин исследования в Центрально-Лесном государственном биосферном природном заповеднике: итоги и перспективы // Научные чтения памяти профессора , вып. 4. Смоленск: Изд-во СГПУ, 2004. С. 127-135. (0,5 п. л., авторский вклад – 50 %)
64. Истомин корреляционного анализа признаков многомерных объектов в мониторинге популяций // Матер. региональной конф. с междунар. участием "Северо-Западная Россия: Проблемы экологии и социально-экономического развития, Псков, 2004. С. 193-198. (0,4 п. л.).
65. Истомин изменений климата и природных катастрофических явлений на биосистемы мелких млекопитающих // Запад России и ближнее зарубежье: устойчивость социально-культурных и эколого-хозяйственных систем социально-экономического развития. Матер. межрегиональной научной конфер. с междунар. участием, Псков, 2005. С. 91-98. (0,5 п. л.).
66. Истомин мониторинг природно-очаговых инфекций // Псковский регионологический журнал, №1, Псков, 2005. С. 122-1п. л.).
67. Истомин итоги использования мелких млекопитающих в биомониторинге лесных экосистем Центрально-Лесного биосферного заповедника // Комплексные исследования в Центрально-Лесном государственном природном биосферном заповеднике: их прошлое, настоящее и будущее. Материалы совещания. Труды Центрально-Лесного заповедника. Вып. 4, Тула, 2007. С. 60-79. (1,3 п. л.).
68. , , Никитина -очаговые инфекции в Центрально-Лесном биосферном государственном заповеднике // Комплексные исследования в Центрально-Лесном государственном природном биосферном заповеднике: их прошлое, настоящее и будущее. Материалы совещания. Труды Центрально-Лесного заповедника. Вып. 4, Тула, 2007. С. 444-461. (1,2 п. л. – авторский вклад – 70%).
69. Истомин численности и фенетического разнообразия популяций мелких млекопитающих в лесных экосистемах Каспийско-Балтийского водораздела Русской равнины // Заповедники России и устойчивое развитие. Матер. конф. Труды Центрально-Лесного государственного природного биосферного заповедника. Вып. 5, Великие Луки, 2007. С. 279-300. (0,8 п. л.).
70. Истомин флуктуации и популяционная динамика ценозообразующих видов в эталонных лесных экосистемах Главного Русского водораздела // Вестник Псковского государственного педагогического университета. Серия естественные и физико-математические науки. Вып. 2. Псков, 2007. С. 45-п. л.).
71. Истомин разнообразие реликтового вида красной сибирской полевки в Центрально-Лесном государственном природном биосферном заповеднике // Северо-Запад России: Эколого-хозяйственные проблемы и перспективы трансграничного сотрудничества. Материалы конференции с международным участием. Псков, 2007. С. 120-124. (0,3 п. л.).
72. Истомин -функциональная организация сообществ и популяций мелких млекопитающих: оценка, диагностика и прогнозирование состояния экосистем // Научные исследования в заповедниках и национальных парках Российской Федерации за гг., вып. 3. Ч. 1., М., 2007. С. 442-443. (0,2 п. л.).
73. Истомин экологической дестабилизации среды на изменчивость и скоррелированность развития признаков // Вестник Псковского государственного педагогического университета. Серия естественные и физико-математические науки. № 4. Псков, 2008. С. 13-23. (1,0 п. л.).
74. Истомин изучение мелких млекопитающих в Центрально-Лесном государственном природном биосферном заповеднике // Центрально-Лесной заповедник – вклад в отечественную и мировую науку. Материалы посвященные 75-летию ЦЛГПБЗ. Великие Луки, 2008. С. 78-85. (0,7 п. л.).
75. Скоррелированность развития морфологических признаков на разных фазах динамики численности популяций // Вестник Псковского государственного педагогического университета. Серия естественные и физико-математические науки. № 6. Псков, 2008. С. 13-23. (1,0 п. л.).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
