

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Фактор транскрипции иммунопрофиля, в большинстве случаев CHL был следующим: В SAP-положительный; Oct-2-отрицательный или BOB.1-отрицательный; и PU.1-отрицательный. Этот фенотип наблюдался в 74% (42/57) случаев CHL. Хотя имеются литературные различия, экспрессия фактор BSAP наблюдалась почти в 90% всех сообщенных случаев CHL (Список 2).
С другой стороны экспрессия фактора Oct-2 и коактиватора BOB.1 наблюдалась в меньшем числе случаев (20% и 34 %, соответственно).
Наши результаты для фактора BSAP в CHL соответствовали большинству публикаций в литературе18,18,22.Исключением явилось изучения Krenacs et al23, показывающие положительные результаты в 5 (36%) из 16 случаев CHL. В отличие от других исследователей, была использована поликлональная подготовка. Интересно, что поликлональные антитела также оценивались Foss et al16, при этом ответ по сравнению с моноклональным был ниже.
Различия в экспрессии факторов Oct-2 и BOB.1 были опубликованы в литературе. Наши результаты приближались к результатам Stein et al, однако в отличие от наших результатов не отмечалась экспрессия сразу двух факторов Oct-2 and BOB. 1. Re et al11, показали отсутствие Oct-2 and BOB.1 во всех изученных случаях CHL. Re et al использовали моноклональные антитела амино-концевых Oct-2, в то время как наши антитела были поликлональными с карбоксильной-концевой молекулой. Saez et al12, также использовали поликлональную подготовку, и обнаруживали факторы Oct-2 и BOB.1 в 71% из 24 случаев CHL с коэкспрессией в большинстве клеток RS, 2х факторов транскрипции. Отмеченные несоответствия могут быть отнесены к небольшому числу проведенных изучений, и различных методов иммуногистохимии.
Факторы транскрипции фенотипа для всех случаев NLPHL и DLBCL в нашем изучении составили BSAP-положительный/Oct-2-положительный/BOB.1-положительный.
Наши результаты в большинстве согласовывались с результатами, опубликованными в литературе16-18: экспрессия фактора BSAP в 100% случаев NLPHL и в большинстве случаев DLBCL. Krenacs et al.23 обнаруживали экспрессию BSAP только в 2х из 4 случаев NLPHL и только в 79% случаев DLBCL, эти результаты могут быть связаны с использованием поликлональных антител. Обнаружение факторов Oct-2 и BOB.1 в NLPHL также согласовывалось с литературой, которая показывала экспрессию факторов Oct-2 и BOB.1 во всех случаях NLPHL10,12,17. Наши результаты для DLBCL (включая TCRLBCL) были несколько выше, однако в литературе было показано большее число случаев экспрессии Oct-2 и BOB.1 во всех изучаемых случаях12,17. Хотя клетки Ходжкина и Рида - Штернберга были отнесены к зародышевым B - клеткам1,2, экспрессия маркеров CD20, CD22, и CD79a, при этом наблюдалась в меньшем числе случаев.
Список 3 вносит результаты лечения различных изучений, которые оценивали экспрессию B- клеточных маркеров в CHL, NLPHL, и DLBCL10,17,18,21,24,25.
В литературе, описаны в случаи CHL: экспрессия CD20 маркера наблюдалась в (23.76% среди всех случаев), далее наличие CD22 составило (14.6%) и CD79a (10.9%). Данные относительно экспрессии CD20 в NLPHL составили (97.0%), с относительно меньшим числом случаев экспрессии CD22 (12/14[86%]) и CD79a (27/33[82%]). Процент обнаружения маркера CD79a, был намного выше в исследованиях при использовании HIER. Отмечалось снижение экспрессии B-клеточных маркеров в DLBCL. По нашим результатам и по литературным источникам, почти во всех случаях, экспрессия CD20 была выше экспрессии CD79a. Оценка экспрессии маркера CD22 в DLBCL не проводилась.
Список 3. Литературные данные экспрессии маркеров CD20, CD22, и CD79a в образцах тканей.
Антитела и использованные клоны | CHL | LPHL | |||||||||||
CD20/ | CD20/ | CD20/ | CD20/ | ||||||||||
Ссылка и год | CD20 | CD22 | CD79a | CD20 | CD22 | CD79a | 22 | 79a | CD20 | CD22 | CD79a | 22 | 79a |
Chittal et al,24 1990 | L26 | 4/4 | |||||||||||
Coles etal,25 1988 | L26 | — | — | 3/25 | — | — | — | — | 15/16 | — | — | — | — |
Hall etal,26,271988 | L26 | — | MB-1 (NS) | 5/38 | — | 0/38 | — | — | 3/3 | — | 1/5 | — | — |
Pinkus and Said,28 1988 | L26 | — | — | 8/63 | — | — | — | — | 9/9 | — | — | — | — |
Schmid etal,29 1991 | L26 | — | 32/55 | — | — | — | — | — | — | — | — | — | |
Zukerberg etal,30 1991 | L26 | — | — | 7/20 | — | — | — | — | — | — | — | — | — |
Carbone et al,31 1992 | L26 | — | — | 11/66 | — | — | — | — | — | — | — | — | — |
Delsol etal,32 1993 | L26 | 4KB128 | HM57 | 17/63 | 7/63 | 0/63 | 3/3 | 0/63 | 3/3 | 3/3 | 3/3 | 3/3 | 3/3 |
Enblad etal,33 1993 | L26 | — | — | 39/151 | — | — | — | — | 1/1 | — | — | — | — |
Kurtin and Roche,34 1993 | L26 | — | — | — | — | — | — | — | — | — | — | — | — |
Kuzu etal,35 1993 | L26 | 4KB128 | HM-57 | 22/79 | 15/79 | 0/79 | NS | NS | 6/7 | 7/7 | 6/7 | NS | 6/7 |
Bai et al,36 1994 | L26 | — | — | 15/64 | — | — | — | — | — | — | — | — | — |
Chadburn and Knowles,37 1994 | L26 | — | — | — | — | — | — | — | — | — | — | — | — |
Korkolopoulou et al,38 1994 | L26 | — | JCB117 | 20/67 | — | 19/94 | — | 7/67 | 7/8 | — | 14/14 | — | 7/8 |
O'Grady et al,39 1994 | L26 | — | — | 7/33 | — | — | — | — | 3/4 | — | — | — | — |
Ashton-Key et al,40 1995 | L26 | — | JCB117 | 8/15 | — | 6/15 | — | 4/15 | 4/4 | — | 3/4 | — | 3/4 |
Mason etal,41 1995 | — | — | JCB117 | — | — | — | — | — | — | — | — | — | — |
Orazi etal,42 1995 | L26 | — | MB-1 (NS) | 9/29 | — | 5/29 | — | 2/29 | — | — | — | — | — |
Herbst etal,43 1996 | L26 | S-HCL1 | — | 48/278 | 10/57 | — | NS | — | 29/31 | 2/4 | — | NS | — |
Vasef etal,44 1997 | L26 | — | NS | 3/43 | — | 2/44 | — | NS | — | — | — | — | — |
Foss etal,16 1999 | L26 | — | — | 8/27 | — | — | — | — | — | — | — | — | — |
Watanabe et al,45 2000 | L26 | — | NS | 18/51 | — | 13/50 | — | 11/50 | — | — | — | — | — |
Stein et al,10 2001 | L26 | — | JCB117 | 6/32 | — | NS | — | — | 35/35 | — | NS | — | — |
Torlakovic et al,17 2001 | NS | — | — | 10/35 | — | — | — | — | 15/15 | — | — | — | — |
Torlakovic etal,18 2002 | L26 | — | — | 25/117 | — | — | — | — | 58/58 | — | — | — | — |
Общее количество | — | — | — | 321/ 1,351 | 29/199 | 45/412 | 3/63 | 23/6324 | 192/198 | 12/14 | 27/33 | 3/3 | 19/22 |
(23.76) | (14.6) | (10.9) | (5) | (10.7) | (970) | (86) | (82) | (100) | (86) | ||||
Настоящее изучение | L26 | FPC1 | HM57 | 17/57 | 10/54 | 3/53 | 6/54 | 2/53 | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 |
CHL, классическая лимфома Ходжкина; DLBCL лимфома из крупных B-клеток; NLPHL узловая лимфома Ходжкина с лимфоцитарным преобладанием; NS, не дифференцируемая.
Схема 1, является алгоритмом для использования факторов транскрипции в сочетании с B- клеточными маркерами в дифференциальном диагнозе лимфомы Ходжкина.
С тех пор как доказали, что BSAP относится к B-клеточному происхождению, приблизительно в 90% случаев, наличие BSAP исключает диагноз ALCL. Однако важно обратить внимание, что недостаточность экспрессии фактора BSAP, не исключает диагноз CHL, так как подтипы CHL в (9%) случаев не экспрессируют BSAP. В нашем изучении фактор Oct-2 был отрицательный во всех случаях ALCL, тогда как BOB.1 был слабо положителен в 2 (25%) из 8 случаев.
При использовании поликлональных антител, совместные результаты Saez et al12 и Marafioti et al46 показали в 15% (2/13) и 77% (10/13), присутствие Oct-2 и BOB.1, соответственно в ALCL. Таким образом, в отличие от BSAP, факторы Oct-2 и BOB.1 не могут использоваться в дифференциальном диагнозе между CHL и ALCL.
Дифференциальный диагноз лимфомы Ходжкина также включает DLBCL.21
В большинстве случаев CHL в нашем исследовании (74% [42/57]), обнаруживался фактор BSAP, при отсутствии в большинстве случаев DLBCL, экспрессии факторов Oct-2 или BOB.1, при экспрессии в большинстве случаев TCRLBCL, всех 3 маркеров. BSAP при этом не помогает в дифференциальной диагностике, так как экспрессируется в большинстве случаев CHL и во всех случаях DLBCL.
Наличие экспрессии сразу 3 маркеров транскрипции, также помогает в установке диагноза, так как в большинстве случаев CHL (10/57 [18%]), экспрессируются все 3 фактора транскрипции.
Однако наблюдается недостаточная экспрессия факторов Oct-2 или BOB.1 в CHL, начиная со всех случаев DLBCL. Хотя, для CHL факторы Oct-2, BOB.1 могут быть положительны, наблюдается экспрессия в слабом диапазоне.
Кроме, того, присутствие всех 3 B - клеточных маркеров говорит в пользу диагноза DLBCL, потому что в большинстве случаев DLBCL (91%), и ни в одном случае CHL, происходит экспрессия все 3 B - клеточных маркеров.
|

Обратите внимание, что основанная на нашем алгоритме, в нашем ряду, фракция всех случаев CHL составляет (7% [4/57]), не отделяя от случаев DLBCL (8% [2/25]), при этом все случаи имеют BSAP-позитивный/Oct-2-позитивный/BOB.1-позитивный фенотип факторов транскрипции, при экспрессии 2 или 3 поверхностных B - клеточных маркеров.
DLBCL и CHL DLBCL/NLPHL |
|
|
|
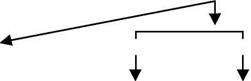
BSAP |
ALCL Oct-2/BOB.1 |
2+ |
3+ |
0/1+ 2 + |
CHL B-клеточные маркеры CD20/CD22/CD79a |
0/1 + |
Схема 1. Предложенный алгоритм дифференциальной диагностики классической лимфомы Ходжкина (CHL) с использованием факторов транскрипции и B - клеточных маркеров. ALCL, анапластическая лимфома из крупных клеток; BOB.1 обязательный B-клеточный белок; специфичный B-клеточный активатор белка BSAP; DLBCL, лимфома из крупных B-клеток; NLPHL, узловая лимфома Ходжкина с лимфоцитарным преобладанием; Oct-2, октаймер - обязательный фактор транскрипции 2; 3+, экспрессия 3 маркеров; 2+, экспрессия 2 маркеров; 1+, экспрессия 1 маркера; нет экспрессии маркеров.
Изменения в транскрипции B - клеток и поверхностных маркеров в случаях CHL, были положены в основу дифференциации B- клеток в CHL.
Примечательно, что в 3х из 4 случаев CHL с лимфоцитарным преобладанием (LRCHL), обнаруживался BSAP-положительный/Oct-2-положительный/BOB.1-положительный фенотип, наблюдаемый и в NLPHL.
Хотя число подтвержденных случаев слишком мало, для того чтобы сделать заключение, это наводит на мысль о том, что подтип LRCHL, связан с NLPHL, ближе, чем другие подтипы в CHL47. Подобный фенотип факторов транскрипции для NLPHL и LRCHL, приводит к поддержанию нормального B - клеточного фенотипа.
Как техническое примечание, мы обнаружили, что с помощью моноклональных антител для BSAP и поликлональных для Oct-2 и BOB.1, легко получались и интерпретировались результаты.
Кроме того, хотя наши результаты, согласовывались с Torlakovic et al17 которые обнаруживали PU.1 положительный, только в 2 (6%) из 35 случаев CHL, мы не смогли подтвердить экспрессию PU. 1 во всех случаях NLPHL.
Наши результаты подтверждают происхождение CHL и NLPHL из B - клеток. Также обнаружение различных B - клеточных факторов транскрипции и B- клеточных маркеров экспрессии в CHL, определяет спектр для дифференцирования B - клеток в пределах этой группы.
Мы обнаружили, что из-за специфичного B - клеточного происхождения в CHL, BSAP является лучшим маркером для дифференциальной диагностики CHL и фенотипа нулевых/ клеток ALCL.
Таким образом, хотя рутинные морфологические и иммуногистохимические изучения помогают в диагностике CHL от других подтипов, экспрессия различных факторов BSAP, Oct-2, and BOB.1 и B - клеточных маркеров CD20, CD22, и CD79a в трудных случаях помогает в диагностике CHL от NLPHL и DLBCL.
Литература.
1. Kuppers R, Rajewsky K, Zhao M, et al. Hodgkin disease: Hodgkin and Reed-Sternberg cells picked from histological sections show clonal immunoglobulin gene rearrangements and appear to be derived from B cells at various stages of development.
Proc Natl Acad Sci USA. 1994;91:.
2. Hummel M, Ziemann K, Lammert H, et al. Hodgkin's disease with monoclonal and polyclonal populations of Reed - Sternberg cells. N EnglJMed. 1995;333:901-906.
3. Kanzler H, Kuppers R, Hansmann ML, et al. Hodgkin and Reed-Sternberg cells in Hodgkin's disease represent the outgrowth of a dominant tumor clone derived from (crippled) germinal center B cells. J Exp Med. 1996;184:.
4. Marafioti T, Hummel M, Foss H-D, et al. Hodgkin and Reed - Sternberg cells represent expansion of a single clone originating from a germinal center B-cell with functional immunoglobulin gene rearrangements but defective immunoglobulin transcription. Blood. 2000;95:.
5. Scholer HR. Octamania: the POU factors in murine development. Trends Genet. 1991;7:323-329.
6. Theil J, Laumen H, Marafioti T, et al. Defective octamer - dependent transcription is responsible for silenced immunoglobulin transcription in Reed-Sternberg cells.
Blood.2001;97:.
7. Latchman DS. The Oct-2 transcription factor. IntJ BiochemCellBiol. 1996;28:.
8. Chang JF, Phillips K, Lundback T, et al. Oct-1 POU and octamer DNA cooperate to recognize the Bob-1 transcription coactivator via induced folding. J Mol Biol. 1999;288:941-952.
9. Laumen H, Nielsen PJ, Wirth T. The BOB.1/OBF.1 coactivator is essential for octamer-dependent transcription in В cells. Em] Immunol. 2000;30:458-469.
10. Stein H, Marafioti T, Foss H-D, et al. Down-regulation of BOB. 1/OBF. 1 in classical Hodgkin disease but not in lymphocyte predominant Hodgkin disease correlates with immunoglobulin transcription. Blood. 2001;97:496-501.
11. Re D, Muschen M, Tahaman A, et al. Oct-2 and Bob-1 deficiency in Hodgkin and Reed-Sternberg cells. Cancer Res. 2001;61:.
12. Saez A-I, Artiga M-J, Sanchez-Beato M, et al. Analysis of the octamer-binding transcription factors Oct2 and Oct1 and their coactivator BOB.1/OBF.1 in lymphomas.
Mod Pathol. 2002;15:211-220.
13. Adams B, Dorfler P, Aguzzi A, et al. Pax-5 encodes the transcription factor BSAP and is expressed in B lymphocytes, the developing CNS, and adult testis.
Genes Dev. 1992;6:.
14. Urbanek P, Wang ZQ, Fetka I, et plete block of early В cell differentiation and altered patterning of the posterior midbrain in mice lacking Pax/BSAP. Cell. 1994;79:901-912.
15. Kozmik Z, Wang S, Dorfler P, et al. The promoter of the CD19 gene is a target for the B-cell-specific transcription factor BSAP. Mol Cell Biol. 1992;12:.
16. Foss H-D, Reusch R, Demel G, et al. Frequent expression of the B-cell—specific activator protein in Reed-Sternberg cells of classical Hodgkin's disease provides further evidence of its B - cell origin. Blood. 1999;94:.
17. Torlakovic E, Tierens A, Dang HD, et al. The transcription factor PU. 1, necessary for B-cell development is expressed in lymphocyte predominance, but not classical Hodgkin's disease.
AmJ Pathol. 2001;159:.
18. Torlakovic E, Torlakovic G, Nguyen PL, et al. The value of anti—PAX-5 immunostaining in routinely fixed and paraffin - embedded sections: a novel pan pre-B and B-cell marker. Am
JSurgPathol. 2002;26:.
19. McKercher SR, Torbett BE, Anderson KL, et al. Targeted disruption of the PU.1 gene results in multiple hematopoietic abnormalities. EMBOJ. 1996;15:.
20. Oikawa T, Yamada T, Kihara-Negishi F, et al. The role of Ets family transcription factor PU. 1 in hematopoietic cell differentiation, proliferation and apoptosis.
Cell Death Differ.1999;6:599-608.
21. Jaffe ES, Harris NL, Stein H, et al, eds. Pathology and Genetics of Tumours of Haematopoietic and Lymphoid Tissues. Lyon, France: IARC Press; 2001. World Health Organization Classification of Tumours.
22. Hertel CB, Zhou X, Hamilton-Dutoit SJ, et al. Loss of B-cell identity correlates with loss of B cell-specific transcription factors in Hodgkin/Reed-Sternberg cells of classical Hodgkin lymphoma. Oncogene. 2002;21:.
23. Krenacs L, Himmelmann AW, Quintanilla-Martinez L, et al. Transcription factor B-cell-specific activator protein (BSAP) is differentially expressed in B cells and in subsets of B-cell
lymphomas. Blood. 1998;92:.
24. Chittal SM, Alard C, Rossi J-F, et al. Further phenotypic evidence that nodular, lymphocyte-predominant Hodgkin's disease is a large B-cell lymphoma in evolution.
AmJ SurgPathol. 1990;14:.
25. Coles FB, Cartun RW, Pastuszak WT. Hodgkin's disease, lymphocyte-predominant type: immunoreactivity with B-cell antibodies. ModPathol. 1988;1:274-278.
26. Hall PA, d'Ardenne AJ, Stansfeld G. Paraffin section immunohistochemistry, I: non-Hodgkin's lymphoma. Histopathology. 1988;13:149-160.
27. Hall PA, d'Ardenne AJ, Stansfeld G. Paraffin section immunohistochemistry, II: Hodgkin's disease and large cells anaplastic (Ki1) lymphoma. Histopathology. 1988;13:161-169.
28. Pinkus GS, Said JW. Hodgkin's disease, lymphocyte predominance type, nodular: further evidence for a B cell derivation: L&H variants of Reed-Sternberg cells express L26, a pan B cell marker. AmJ Pathol. 1988;133:211-217
29. Schmid C, Pan L, Diss T, et al. Expression of B-cell antigens by Hodgkin's and Reed-Sternberg cells. AmJ Pathol.1991;139:701-707.
30. Zukerberg LR, Collins AB, Ferry JA, et al. Coexpression of CD15 and CD20 by Reed-Sternberg cells in Hodgkin's disease. AmJ Pathol. 1991;139:475-483.
31. Carbone A, Gloghini A, Volpe R. Paraffin section immunohistochemistry in the diagnosis of Hodgkin's disease and anaplastic large cell (CD30+) lymphomas.
Virchows Arch A Pathol Anat Histopathol. 1992;420:527-532.
32. Delsol G, Meggeto F, Brousset P, et al. Relation of follicular dendritic reticulum cells to Reed-Sternberg cells of Hodgkin's disease with emphasis on the expression of CD21 antigen. Am
] Pathol. 1993;142:.
33. Enblad G, Sundstrom C, Glimelius B. Immunohistochemical characteristics of Hodgkin and Reed-Sternberg cells in relation to age and clinical outcome.
Histopathology.1993;22:535-541.
34. Kurtin PJ, Roche PC. Immunoperoxidase staining on non - Hodgkin's lymphomas for T-cell lineage associated antigens in paraffin sections: comparison of the performance characteristics of four commercially available antibody preparations. Am J Surg Pathol. 1993; 17:898-904.
35. Kuzu I, Delsol G, Jones M, et al. Expression of the Ig - associated heterodimer (mb-1 and B29) in Hodgkin's disease. Histopathology. 1993;22:141-144.
36. Bai MC, Jiwa NM, Horstman A, et al. Decreased expression of cellular markers in Epstein-Barr virus—positive Hodgkin's disease. J Pathol. 1994;174:49-55.
37. Chadburn A, Knowles DM. Paraffin-resistant antigens detectable by antibodies L26 and polyclonal CD3 predict the B - or T-cell lineage of 95% of diffuse aggressive non- Hodgkin's lymphomas. Am J Clin Pathol. 1994;102:284-291.
38. Korkolopoulou P, Cordell J, Jones M, et al. The expression of the B-cell marker mb-1 (CD79a) in Hodgkin's disease. Histopathology. 1994;24:511-515.
39. O'Grady JT, Stewart S, Lowrey J, et al. CD40 expression in Hodgkin's disease.
AmJ Pathol. 1994;144;21-26.
40. Ashton-Key M, Thorpe PA, Allen JP, et al. Follicular Hodgkin's disease.
AmJ SurgPathol. 1995;19:.
41. Mason DY, Cordell JL, Brown MH. CD79a: a novel marker for B-cell neoplasms in routinely processed tissue samples. Blood. 1995;86:.
42. Orazi A, Jiang B, Lee CH, et al. Correlation between presence of clonal rearrangements of immunoglobulin heavy chain genes and B-cell antigen expression in Hodgkin's disease. Am
JClinPathol. 1995;104:413-418.
43. Herbst H, Raff T, Stein H. Phenotypic modulation of Hodgkin and Reed-Sternberg cells by Epstein-Barr virus. JPathol. 1996; 179:54-59.
44. Vasef MA, Alsabeh R, Medeiros LJ, et al. Immunophenotype of Reed-Sternberg and Hodgkin's cells in sequential biopsy specimens of Hodgkin's disease: a paraffin-section immunohistochemical study using the heat-induced epitope retrieval method.
AmJ Clin Pathol. 1997; 108:54-59.
45. Watanabe K, Yamashita Y, Nakayama A, et al. Varied B-cell immunophenotype of Hodgkin/Reed-Sternberg cells in classic Hodgkin's disease. Histopathology. 2000;36:353-361.
46. Marafioti T, Ascani S, Pulford K, et al. Expression of B - lymphocyte-associated transcription factors in human T-cell neoplasms. AmJ Pathol. 2003;162:861-871.
47. Anagnostopoulos I, Hansmann ML, Franssila K, et al. European Task Force on Lymphoma project on lymphocyte predominance Hodgkin disease: histologic and immunohistologic analysis of submitted cases reveals 2 types of Hodgkin disease with a nodular growth pattern and abundant lymphocytes. Blood. 2000; 96:

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 |
