


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
элемент | Са | К | N |
листья | 47 | 44 | 111 |
ветви | 63 | 20 | 75 |
Подсчитано, что за счет выборочной рубки леса, прореживания и других примеров использования богатств леса человеком почва в лесах истощается все же меньше и не так быстро, как при возделывании сельскохозяйственных культур. Однако, некоторые запрещенные сейчас способы хозяйствования могли разомкнуть циклы биогенных элементов и вызвать значительное обеднение лесных экосистем. К таковым относятся: выкашивание травы в лесу; выпас скота; сбор опавших листьев на подстилку для скота; корчевание пней и выжигание растительности с целью использования золы в качестве удобрения под сельскохозяйственные культуры; снятие дерна и сгребание подстилки для использования в качестве удобрения и т. д.
ГЛАВА 5. ЭНЕРГИЯ В ЭКОЛОГИЧЕСКИХ
СИСТЕМАХ
Все разнообразие различных проявлений жизни сопровождается превращениями энергии. Единственным источником энергии для биосферы является Солнце. Энергия, получаемая в виде света поверхностью Земли, уравновешивается энергией, излучаемой с ее поверхности в форме невидимого теплового излучения. Это сохраняет тепловой баланс Земли на постоянном уровне. Данные климатологии не дают оснований говорить о сколько-нибудь заметном потеплении или похолодании Земли и ее атмосферы. Таким образом осуществляется круговорот энергии на Земле как космическом объекте.
Иной вид имеет передача и превращение энергии в пределах биосферы, экосистем, в частности. Сущность жизни состоит в непрерывной последовательности таких изменений, как рост, самовоспроизведение и синтез сложных химических соединений. Все разнообразие проявлений жизни сопровождается превращением энергии, хотя энергия при этом не создается и не уничтожается (первый закон термодинамики). Без переноса энергии не было бы ни жизни, ни экологических систем. Мы, разумные существа, не должны забывать, что наша цивилизация - лишь одно из замечательных явлений природы, зависящих от постоянного притока концентрированной энергии светового излучения.
На верхнюю границу биосферы из космоса падает солнечный свет с энергией 8,3 Дж на 1 см2 в 1 мин, но, проходя через атмосферу, он экспоненциально ослабляется, и в ясный летний полдень до поверхности Земли может дойти не более 67 % энергии. Растения умеренной зоны получают за день энергии в среднем 125кДж/м2. Из падающей на автотрофный компонент биосферы - на продуценты - световой энергии, лишь очень небольшая часть поглощается зелеными растениями и превращается в тепло, покидающее затем и растение, и биосферу. Весь животный мир получает эту потенциальную химическую энергию из органических веществ, созданных фотосинтезирующими растениями. Например, животные поглощают химическую потенциальную энергию пищи и большую часть ее превращают в тепло за счет процессов окисления пищи в организме. Тепло необходимо для пополнения больших энергетических затрат животных организмов, например, для движения с целью поиска пищи и др. Меньшая же часть энергии пищи у животных переводится также в химическую потенциальную энергию заново синтезированной протоплазмы с накоплением биомассы соответствующего уровня потребления.
Биомассой экосистемы называют совокупность организмов, растительных, животных, почвенных, микроорганизмов, присутствующих в экосистеме в момент наблюдения. Она может быть выражена числом особей, в весовых и энергетических единицах (джоулях).
Продуктивность - это скорость продуцирования биомассы.
Первичной продуктивностью называют скорость, с которой продуценты (зеленые растения) в процессе фотосинтеза связывают энергию и запасают ее в форме органических веществ. Эти вещества могут быть использованы растительноядными организмами, то есть консументами, в качестве пищи.
Вторичная продуктивность означает биомассу, продуцируемую консументами или организмами, разлагающими отмершую материю.
5.1. Поток энергии в экосистеме
В экосистемах различают поток энергии на уровне продуцентов (зеленых растений) и на уровне консументов (животных).
Исходя из приведенного определения продуктивности,
потоком энергии на уровне продуцентов называют
первичную валовую продуктивность - ПВ, представляющую собой суммарную продукцию фотосинтеза, включающую чистую продуктивность (Пч) и вещество, расходуемое на дыхание (Д) за период наблюдения, т. е. Пв=Пч+Д.
Чистая первичная продуктивность (Пч) при этом есть видимый фотосинтез. Это скорость накопления создаваемого органического вещества сверх того, которое затрачено на дыхание, т. е. Пч = Пв - Д. Иными словами Пч - это вещество, которое можно взвесить при уборке урожая.
Поток энергии для любого уровня потребления (травоядные, хищники и др.) представляет собой суммарную ассимиляцию (А) на этом уровне, или это сумма энергии потенциальной, заключенной в продуцированной этим уровнем биомассе и энергии веществ, затраченных на дыхание (Д) в процессе продуцирования биомассы. Эта биомасса есть вторичная продуктивность (П), согласно определению, данному выше.
Таким образом, поток энергии для травоядных выразится как А1 = П1 + Д1 - первый уровень потребления;
для хищников - А2 = П2 + Д2 - второй уровень потребления и т. д.
Поток энергии - важное понятие в экологии.
Особенность потока энергии состоит в том, что энергия используется данными организмами (растительными или животными) лишь однократно. Если биогенные элементы, из которых состоят живые вещества, обращаются многократно между живыми и неживыми компонентами экосистемы, то энергия в итоге превращается в организме в тепло и быстро исчезает из экосистемы. Жизнь, таким образом, поддерживается непрерывным потоком солнечного света извне.
Односторонний приток энергии в биосферу есть универсальное явление природы, и представляет собой результат действия законов термодинамики.
Если вспомнить первый закон термодинамики, то он утверждает, что энергия может переходить из одного вида в другой, например, энергия света переходит в потенциальную энергию синтезированных органических веществ. Второй закон термодинамики гласит, что не может быть ни одного процесса, связанного с превращением энергии, без деградации ее из концентрированной формы в рассеянную. То есть, не может быть 100 %-го превращения энергии света в пищу и пищи в тепло.
Можно утверждать, что односторонний поток энергии и круговорот веществ являются двумя великими принципами или законами общей биологии.
5.2. Эффективность потока энергии
Эффективность передачи потока энергии удобно проследить на примере простой пищевой цепи: продуценты, травоядные, хищники (рис.4).
На рис.4 кормовая цепь представлена : слева (условно) - зелеными растениями, затем травоядными - зайцем и лисой - хищником.
Из общего поступления энергии с солнечным светом С половина, С1, поглощается растениями, а вторая половина света, падающего на растения, рассеивается в виде тепла. Поток энергии для продуцентов обозначен Пв. Часть валовой продукции (Пв) расходуется на дыхание растений Д, а другая ее часть - на создание чистой продукции Пч. В свою очередь часть чистой продукции Пч служит кормом К1 растительноядным. Остальная часть Пч остается неиспользованной Н, она отмирает и поступает в почву в пищу биоредуцентам - разлагателям.
Поток энергии в экосистеме
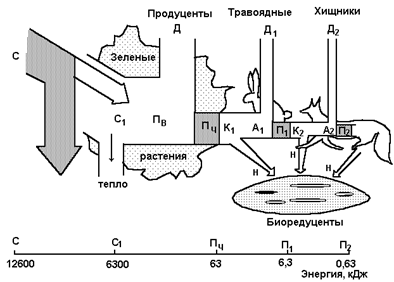
Рис. 4
ПВ - валовая продукция; Пч - чистая продукция; А1, А2 - ассимилированная часть корма; П1, П2 - вторичная продуктивность травоядных и хищников; Д - энергия, затраченная на дыхание; Н - неиспользованная часть корма.
Из величины К1, съеденной растительноядными, ассимилируется только некоторое количество А1, причем из этого А1 часть идет на создание биомассы П1, а для создания биомассы используется энергия второй части ассимилированного корма, выделяемая при дыхании уровня травоядных Д1. Следовательно, вторичная продуктивность растительноядных имеет вид:
П1=А1-Д1,
а поток энергии, проходящий через первый уровень потребления, выражается:
А1=П1+Д1
Мы подходим, таким образом, к трофическому уровню плотоядных, т. е. хищников, консументов 2-ого порядка.
Хищники не истребляют всех возможных жертв, а из той доли, которую они потребляют, в качестве корма К2 ассимилируют часть вещества, используя его на создание биомассы этого уровня П2; другая часть затрачивается на дыхательную энергию Д2. Поток энергии, проходящий через трофический уровень плотоядных, будет: А2 = П2 + Д2.
Под рисунком приведена шкала энергии для каждого уровня, начиная с поступления в экосистему солнечного излучения в количестве 12600 кДж/м2 в день для умеренной зоны.
Из шкалы следует, что из половины ассимилированной солнечной энергии С1 = 6300 кДж в энергию пищи (Пч) превращается 1% энергии, 63 кДж. Эта величина, 1 %, есть среднее значение эффективности видимого фотосинтеза. Как видно, эта эффективность в природе очень низка. Вторичная продуктивность уровней потребления (П1 и П2) составляет 10 % предыдущей, т. е. на последующий уровень передается около 10 % энергии. Это и есть среднее значение эффективности потока энергии в природе по цепям питания, она также невысока, а ее численное значение называют правилом десяти процентов. Это значит, что биомасса уровня хищников будет меньше биомассы травоядных в 10 раз, а растений в 100 раз, чем собственно и определяется, в частности, численность популяции.
5.3. Экологические пирамиды
Эффективность передачи энергии или экологическая эффективность природной экосистемы определяется отношением величины ассимиляции трофической цепи на данном уровне к величине ассимиляции на предыдущем уровне и выражается в процентах. В принятых обозначениях примера рис.4 эффективность 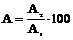 . Для продуцентов - это отношение Пв/С1. Эта эффективность, как было показано, очень низка.
. Для продуцентов - это отношение Пв/С1. Эта эффективность, как было показано, очень низка.
Для наглядности экологическую эффективность продуктивности уровней потребления в экосистеме выражают в виде экологических пирамид, в которых прямоугольники расположены один над другим. Длина же каждого прямоугольника пропорциональна продуктивности каждого уровня (рис.5).
Экологическая пирамида
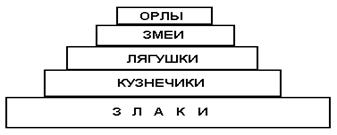
Рис. 5
На рисунке приведен пример простой экосистемы, в которой первичная продукция обеспечивается злаками, а далее идут последовательно уровни потребления.
Высота пирамиды соответствует длине пищевой цепи. Чем длиннее цепь, тем меньше значение по биомассе, числу или энергии имеют плотоядные на вершине пирамиды. Последнее можно наглядно проиллюстрировать пирамидами чисел, биомасс и энергии для очень простой и экологически интересной для анализа пищевой цепи человека.
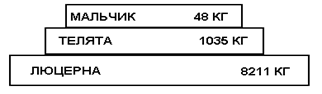
В этом примере исходной продукцией экосистемы принят посев люцерны на площади 4 га. На этом поле кормятся телята, а телятами, в свою очередь, питается консумент 2-го порядка, 12-летний мальчик, в течение года, при этом телятина служит ему единственным источником питания.
Пирамида чисел выглядела бы так:
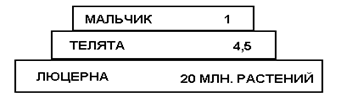
Пирамида биомасс для этого же случая:
Пирамида энергии, включая солнечную, в Дж:
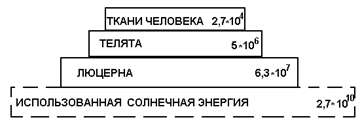
Во всех пирамидах заметно снижение калорийности по мере перехода от нижних уровней к верхним.
Этот теоретический пример хорошо иллюстрирует эффективность различных трофических уровней.
Из солнечной энергии люцерна использует 0,24 %. Из энергии, аккумулированной люцерной, для накопления продукции телят в течение года используется 8 %. и этот коэффициент еще считается высоким, т. к. люцерна считается идеальным кормом, не имеющим больших отходов. Из энергии, аккумулированной телятами, используется на развитие и рост ребенка в течение года всего 0,54 %. Коэффициент использования здесь очень мал, что, помимо других причин, объясняется несъедобностью значительной части биомассы телят.
Низкая способность фотосинтеза при трансформации солнечной энергии в растительное вещество представляет. как отмечено выше, общее природное явление. Снижение коэффициента полезного действия при переходе к другим звеньям кормовой цепи и в итоге к плотоядным выявляется особенно разительно. В конечном счете, из солнечной энергии, падающей на 4 га почвы, используется лишь чуть более одной миллионной доли на пропитание ребенка, а остальная энергия теряется.
Этот наглядный пример показывает как малоэффективно человек использует продукцию биосферы, если находится в конце трофической цепи.
Экологически грамотно было бы пользоваться первичной продукцией, быть ближе к ней, а не расходовать ее на выращивание вторичной, которая идет затем в пищу.
ГЛАВА 6. РАЗВИТИЕ И ЭВОЛЮЦИЯ
ЭКОЛОГИЧЕСКИХ СИСТЕМ
Развитие экосистемы чаще всего называют экологической сукцессией, это процесс последовательной смены природных сообществ.
Признаки сукцессии: 1) сукцессия - это упорядоченныйпроцесс развития сообщества, связанный с изменением во времени видовой структуры сообщества и с изменением протекающих в сообществе процессов; 2) сукцессия происходит в результате изменения физической среды под действием самого же развивающегося сообщества, т. е. сукцессия контролируется сообществом. Изменяющаяся среда определяет при этом характер или направленность сукцессии, скорость развития экосистемы, устанавливает пределы этого развития; 3) сукцессия протекает под влиянием внешних факторов: влажности, температуры, количества осадков, состава почвы, солнечной радиации и др.; 4) кульминацией развития является стабилизированная экосистема, в которой на единицу потока энергии приходится максимальная биомасса, максимально возможное количество видов ипопуляций растительного и животного мира, а также связей между организмами - пищевых, борьбы за пространство и др. Стабилизированную экосистему называют еще зрелой или климаксной экосистемой.
Так как в процессе сукцессии происходит смена сообществ, то переходные сообщества называют последовательными стадиями, стадиями развития или стадиями первых поселенцев.
Замещение видов вызывается тем, что каждая популяция меняет окружающую среду и создает при этом условия, благоприятные для других популяций, которые постепенно сменяют предыдущие. Это происходит до тех пор, пока не будет достигнуто равновесие между биотическими и абиотическими компонентами среды. Развитие экосистем во многом аналогично развитию отдельного организма.
6.1. Виды сукцессии
Различают первичную и вторичную сукцессии.
Первичная сукцессия начинается на участке, который перед этим не был занят каким-либо сообществом, например, на только что вышедшей над поверхностью воды скале, на песке или лавовом потоке.
Вторичная сукцессия происходит на площади, с которой удалено предыдущее сообщество, например, на заброшенном поле или на вырубке леса. Вторичная сукцессия протекает обычно быстрее, так как на территории, которая ранее была занята, уже имеется некоторые организмы или их зачатки и она более благоприятна для развития сообщества, чем так называемая стерильная зона.
Характерным примером развития первичной сукцессии являются дюны озера Мичиган. Озеро Мичиган было некогда гораздо больше, чем теперь. Отступая к своим современным границам, оно оставляло за собой все более и более молодые песчаные дюны. Это позволяет, переходя от современных дюн до первых возникающих тысячи лет назад, проследить первичную сукцессию (рис.6).
Сообщества первых поселенцев на дюнах состоят из злаков, ивняка, вишни, хлопкового дерева и таких животных, как жуки-скакуны, норные пауки и кузнечики. За сообществами первых поселенцев следуют лесные сообщества, каждое из которых обладает своим животным миром. Несмотря на то, что развитие началось в очень сухом и бесплодном местообитании, оно в конечном счете приводит к буково-кленовому лесу, влажному и холодному, в противоположность голым дюнам. мощная, богатая гумусом почва с дождевыми червями и моллюсками контрастирует с сухим песком, на котором сообщество образовалось. С использованием современных радиоуглеродных методов определения возраста установлено, что для достижения зрелого лесного сообщества на дюнах озера Мичиган требуется около 1000 лет.
Первичная сукцессия
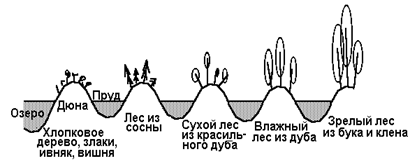
Рис. 6
В качестве примера вторичной сукцессии можно рассмотреть заброшенные пахотные земли, на которых выращивался хлопчатник, кукуруза и др. (рис.7). Характер сукцессии ясен из рисунка.
Вторичная сукцессия
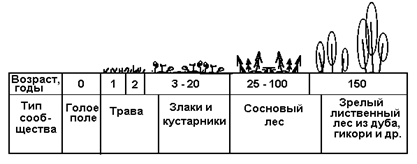
Рис. 7
Длительность ее - 150-200 лет.
Вторичная растительная сукцессия в степных районах так же поразительна, как лесная, только в ней участвуют травянистые растения. Например, это сукцессия на заброшенных проселочных дорогах. Сукцессия слагается из четырех последовательных стадий: 1) стадия однолетних сорняков (2-5 лет); 2) стадия короткоживущих злаков (3-10 лет); 3) стадия многолетних злаков (10-20 лет); 4) стадия зрелых злаков (достигается за 20-40 лет). Природе, таким образом, требуется от 20 до 40 лет, чтобы, начав на голом или изборожденном колеями грунте, “построить” зрелое степное сообщество. Конечные сроки зависят от лимитирующих факторов, таких, как влажность, выпас скота и т. п. Несколько засушливых лет подряд или чрезмерное выедание вызывают возврат сукцессии к стадии однолетних сорняков; насколько далеко может зайти такой возврат, зависит от силы воздействия.
6.2. Сукцессия в водной среде
В водных местообитаниях сукцессию можно наблюдать так же, как и в наземных. Особенно нагляден этот процесс в искусственно создаваемых водоемах - прудах, водохранилищах, подпруженных озерах. При заполнении чаши водоема под воду уходит большое количество органического вещества - это почва с растительностью.
Первой стадией развития водоема является высокопродуктивная стадия “цветения”, для которой характерны интенсивное разложение, высокая активность микроорганизмов, обилие питательных веществ, низкое содержание кислорода в придонном слое воды, т. к. он расходуется на разложение и часто быстрый и энергичный рост рыбы. На этой стадии рыболовы очень счастливы.
Вторая стадия характеризуется стабилизацией водохранилища; ее условно можно назвать зрелой стадией развития экосистемы. Наступает она, когда израсходуется запас питательных веществ и водохранилище стабилизируется на некоторой пониженной продуктивности. Содержание кислорода в придонном слое повышается, так как “цветение” кончилось и расход кислорода на разложение органического вещества уменьшился. Урожай рыбы - низкий, огорчение рыболовам. Эта стадия может длиться очень долго.
Третья стадия - стадия переходных состояний - наступает, когда в результате эрозии почвы в водоем поступает большое количество биогенных элементов, и повторяются вышеописанные процессы. Такие серии смены состояний водной среды могут происходить неоднократно, пока водоем не окажется занесенным почвой. В этом случае сукцессия для водной биоты пошла вспять и закончилась “трагически”, т. е. сильный приток энергии на первой стадии ускорил сукцессию, а затем остановил ее. На этом месте начинается наземная сукцессия. Так происходит с прудами между дюнами оз. Мичиган.
Если водохранилища создаются на истощенных водосборах или на первично стерильных участках, они дают обратную картину сукцессии - низкую продуктивность вначале, которая может повыситься за счет эрозии в водоем. В последующем же процесс развития будет зависеть от поступления веществ, определяемого внешними факторами и экосистемой.
Сукцессия в океане до последнего времени не интересовала ученых, так как океан находится в целом в стационарном состоянии, химически и биологически он застабилизировался на протяжении многих столетий. Однако, по мере тог как в результате загрязнений равновесие в океане начнет нарушаться, сукцессионные изменения в океане привлекают внимание ученых. Эти изменения можно суммировать следующим образом:
возрастает средний размер клеток, что ведет к уменьшению числа особей за счет их укрупнения;
продуктивность или скорость деления снижается;
изменяется химический состав фитопланктона (мельчайших водорослей - продуцентов океана);
в составе зоопланктона наблюдается сдвиг от пассивных фильтратов (организмов, отделяющих пищу от воды фильтрованием) к подвижным организмам - охотникам; этот сдвиг есть ответ на пункт 1, так как уменьшилось число мелких частиц пищи за счет более крупных, ставших более редкими.
6.3. Изменение составляющих потока
энергии в процессе сукцессии
На ранних стадиях экологической сукцессии, когда “природа молода”, уровень первичной продукции или общий (валовой) фотосинтез (Пв) превосходит уровень дыхания сообщества (Д), т. е. в развивающихся системах часть энергии остается. За ее счет образуются различные структуры экосистемы - животный мир, мир микроорганизмов и др., увеличиваются ресурсы данного место обитания.
В зрелых же экосистемах вся фиксированная энергия используется в общем дыхании растений и организмов. Уровень дыхания Д приближается к валовой продукции, а, следовательно. величина чистой продукции падает до нуля, что следует из приведенного выше определения потока энергии - Пв = Пч + Д; при Пв = Д Пч = О. Это значит, что в зрелой экосистеме нет лишней энергии и нет прироста вещества за год.
Отношение Д/Пв свидетельствует о степени приближения экосистемы к состоянию зрелости.
В качестве примера приведем значения Д, Пв, Пч, в г/м2·день, для одной из изученных экосистем сосно-дубового леса, в которой Пв = 2650, Д =2100 и Пч = 650. Величина отношения Д/Пв=2100/2650 = 0,85, или 85 %, показывает, что рассматриваемая лесная экосистема близка к зрелому состоянию с небольшой чистой продукцией.
Пока Пв больше Д, в развивающейся экосистеме будут накапливаться органическое вещество и биомасса (Б).
Взаимные изменения этих величин в процессе сукцессии представлены наглядно на рис.8.
Заштрихованная область показывает, как убывает чистая продукция по мере приближения экосистемы к зрелому состоянию, к этому времени рост биомассы стабилизируется, максмум чистой продукции приходится на возраст экосистемы 30 лет.
Изменение составляющих продуктивности
в процессе сукцессии
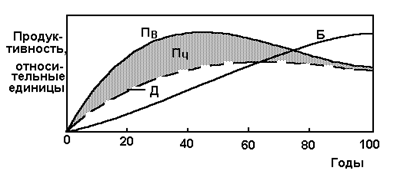
Рис. 8
6.4. Значение экологической сукцессии
Принципы экологической сукцессии чрезвычайно важны для человека.
Зрелое сообщество способно стабилизировать в своем окружении физические факторы среды, и с эволюционной точки зрения это весьма важно, так как оно является буфером на всякие внешние изменения, в том числе и неблагоприятные: бури, наводнения, изменения температуры, ветровую эрозию почвы. Эти функции зрелое сообщество выполняет лучше, чем молодое.
Однако в зрелом сообществе чистая продукция практически отсутствует и, следовательно, использовать такую экосистему с целью получения продуктов питания, заготовки древесины, выпаса скота и др. нельзя. Это равносильно расхищению основного капитала, с которого идут проценты.
Для своих хозяйственных целей, получения продуктов питания, в частности, человек должен располагать ранними стадиями сукцессии, т. е. молодыми сообществами, у которых чистая продукция максимальна. К таким молодым экосистемам относятся возделываемые сельскохозяйственные культуры. Они не достигают состояния зрелости в эволюционном понимании термина “зрелость” - как последняя стадия сукцессии. Особенность этих экосистем состоит в том, что они поддерживаются в требуемом состоянии усилием человека ценой больших вложений дополнительной энергии, затрачиваемой на обработку земли, орошение, производство и внесение удобрений, селекцию, борьбу с вредными насекомыми. В горючем, которое расходуется сельскохозяйственными машинами, заключено не меньше энергии, чем в солнечных лучах, падающих на поле. Если же эти культурные экосистемы предоставить самим себе, то они быстро будут сменены в процессе сукцессии другими природными сообществами. Культурные экосистемы, таким образом, - это неустойчивые экосистемы.
В интересах человека необходимо уделять постоянное внимание обоим видам систем, зрелым и молодым. Если, например, зрелые лесные экосистемы будут уничтожены в погоне за временным доходом от древесины, почва подвергнется водной и ветровой эрозии, продуктивность уменьшится. В итоге очень часто оказывается, что непосредственная выгода меньше, чем та, которая проявилась бы при сохранении лесной экосистемы.
Как же тут быть? Как сохранить зрелые лесные экосистемы и одновременно получить лесную продукцию? Проблема труднейшая. Лесопромышленники решают ее просто и экологически неверно. Они одним махом выбирают всю биомассу, накопленную порой за столетия. Лесосеки же в процессе сукцессии возобновляются менее ценными породами, очень часто задерняются и заболачиваются; нужные человеку леса появляются лишь через столетия. То есть человек сейчас выбирает продуктивность, жертвуя стабильностью. Это очень опасно и уже не раз ввергало человечество в большие беды. Разрушая экологический баланс, человек лишался продукции. Выход есть и, на основании учета принципов экологической сукцессии, требуется постоянный компромисс.
Рациональное лесопользование, например, должно предусматривать не сплошные рубки на тысячах гектаров, а рубку отдельных деревьев или их групп в возрасте хозяйственной спелости. Убрав их, мы получаем пространство, в котором начинают быстро развиваться новые растения. Продуктивность очень велика, мы как бы подстегнули ее вторичной сукцессией, протекающей в окружении зрелой экосистемы, защищающей развивающуюся от неблагоприятных внешних факторов. В итоге мы будем систематически, а не один раз в сто лет получать хозяйственную продукцию, повышать продуктивность сообщества и сохранять его устойчивость. Технически это сложно и дорого, большие капиталовложения окупятся не скоро, но все-таки окупятся. Экологически это единственно возможный путь эксплуатации лесов. Не случайно передовые лесохозяйственники выдвинули сейчас лозунг “Даешь вечные леспромхозы”.
Это пример одной из стратегий хозяйственной деятельности человека - стратегии компромисса между молодыми и зрелыми экосистемами.
Другая стратегия получения продукции и сохранения стабильности экосистем - это стратегия расчленения. Вот что говорит Ю. Одум - один из известных ученых-экологов - по этому поводу: “... было бы чрезвычайно опасно превратить нашу биосферу в один обширный ковер пахотных земель, отметим мимоходом, что эстетический мир, устроенный так, был бы очень скучен. Для нашей собственной безопасности и защиты определенные куски ландшафта должны быть представлены более или менее естественными сообществами. До сих пор не установлено, какой процент площади должны они занимать в различных климатических зонах. Мы только знаем, что когда человек думает о продуктивности, он идет на крайности в обнажении ландшафта.
Остатки цивилизаций и пустыни, возникновение которых обязано деятельности человека, являются прекрасным доказательством того, что человек никогда не осознавал свою гетеротрофную природу и необходимость приспособиться к природе, которой он пытался командовать. Во многих случаях нехватка воды была последней причиной катастроф в жизни человечества”[1].
Однако, как отмечено выше, возделываемые монокультуры тропиков, интенсивная сельскохозяйственная деятельность часто образуют всепожирающий фронт. Истощенные и разрушенные почвы забрасывают, чтобы распахать новые, более плодородные, которые также забрасываются из-за их деградации. За время своей жизни на Земле человек уничтожил не меньше возделываемых земель, чем их существует в настоящее время. Страны, некогда обладавшие плодородными почвами и покрытые лесами, стали теперь безлесыми, превратились в бесплодные территории, неспособные прокормить народы, которые их населяли.
Так, ныне пустынные пространства Ближнего Востока располагали раньше плодородными и лесистыми участками. Сирия, например, снабжала Египет лесом, а Рим маслом и вином. некогда процветавшая Греция, славившаяся зелеными горами и достигавшая непревзойденного уровня цивилизации, ныне заурядная страна с лишенными растительности холмами и каменистыми, чрезвычайно бедными почвами.
Территория Сахары была в период одомашнивания диких животных плодородной областью. Ссылаясь на мнение археологов, С. Боголюбский - крупный профессор-морфолог - пишет, что превращению Сахары в пустыню способствовали стада домашних животных. Они уничтожили растительный покров и выпустили из бутылки джина пустынь. Такой парадокс нередок в истории человечества - внешне целесообразное действие повлекло за собой катастрофические последствия, цветущая область превратилась в одну из крупнейших на Земле пустынь. Сейчас уже 43 % суши - это пустыни и полупустыни, под угрозой опустынивания находится еще 19 % суши (30 млн. км2). Суша деградирует со скоростью 58250 км2/год. Примеры и факты, подобные изложенным, многочисленны.
Стратегия расчленения должна предусматривать такой план ведения хозяйства, при котором можно было бы на разных участках поддерживать интенсивное производство зерна, например, и совершенно нетронутую природу, которая стабилизирует в своем окружении экосистемы и в итоге дает возможность человеку получать продукцию.
Некоторые ученые полагают, что крайне рискованно выводить в хозяйственной деятельности из естественного равновесия более 2./3 территории, занятой природной растительностью. Это максимум, к которому желательно не приближаться.
6.5. Гомеостаз экологических систем
Экосистемы, как и организмы, способны к самоподдержанию и саморегулированию. Анализ этой способности в эпоху интенсивной хозяйственной деятельности человека имеет важное приложение в экологии, поскольку человек все сильнее нарушает природные механизмы контроля состояния среды или пытается заменять естественные механизмы искусственными.
Гомеостаз (от греч. гомео - тот же, стасис - состояние) - термин, применяемый обычно для обозначения способности биологических систем противостоять изменениям и сохранять состояние равновесия.
Модель управления в экологической системе подобна принципам кибернетики в технических системах управления. Например, в обычной системе регулирования температуры в помещении термостат представляет собой датчик с регулятором, исполнительный элемент, а комнатная температура является управляемой величиной. Управление зависит от обратной связи, которая осуществляется, когда сигналы с выхода полностью или частично поступают на вход, при этом, если температура повысилась, то уменьшается мощность нагревателя и наоборот. Когда эта обратная связь положительна, значение температуры возрастает, т. к. регулятор будет давать команду на увеличение мощности нагревателя. Положительная обратная связь - это связь, усиливающая отклонение, необходимая для роста и выживания организмов. Однако для достижения управления, когда надо избежать, например, катастрофического разрастания популяции, должна существовать отрицательная обратная связь. Механизмы, управляющие живыми системами, называют гомеостатическими механизмами. Они многочисленны и разнообразны.
В природе, например, численность любой популяции регулируется в таких пределах, чтобы избежать перенаселения экосистемы. Так, медицинская теория стресса объясняет, что в условиях перенаселенности у высших позвоночных происходит увеличение надпочечников, это один из симптомов сдвига нейроэндокринного равновесия, который в свою очередь сказывается на поведении животных, устойчивости к заболеваниям и другим стрессовым воздействиям. Комплекс таких изменений нередко вызывает стремительное падение плотности популяции. Зайцы-беляки при максимальной плотности часто гибнут от “шока”, связанного с увеличением надпочечников и другими признаками нарушения эндрокринного равновесия. Это пример биологического регулирования численности на уровне популяции.
По одной из теорий регулирование численности популяции осуществляется на уровне экосистемы и объясняется “гипотезой возврата питательных веществ”. Численность леммингов в тундре зависит от качества пищи, а последнее от численности леммингов, в чем и проявляется эффект положительной и отрицательной обратной связи. В результате сильного объедания растительности леммингами в годы взрыва их численности происходит уменьшение в почве доступных минеральных питательных веществ (особенно фосфора) в последующие годы, и питательная ценность пищи леммингов сильно уменьшается. В связи с этим рост и выживание молодых особей значительно снижаются. В течение третьего и четвертого года питательные вещества возвращаются в цикл, растительный покров восстанавливается, и экосистема вновь может обеспечить высокую плотность этих животных.
В больших, зрелых экосистемах поддерживается самокорректирующийся гомеостаз в результате взаимодействия круговорота веществ и потока энергии (см. выше), в связи с чем экосистемы Земли и сама биосфера находятся в устойчивом состоянии. Однако устойчивость экосистем и действие гомеостатических механизмов имеет предел, по достижении которого усиливающиеся обратные связи приводят к гибели системы. В настоящее время человек начинает ускорять процессы в природе, в частности, процессы разложения. Сжигая органическое вещество, запасенное в виде ископаемых горючих веществ (угля, нефти, газа), человек увеличивает поступление углекислоты в атмосферу. Хотя это количество СО2 пока невелико по сравнению с общим его количеством, участвующим в природном круговороте, оно начинает оказывать ощутимый эффект, так как резервуар атмосферы не так уж велик, а океанический резервуар, хотя и больший по объему, не успевает поглощать все новые количества СО2, производимые человеком. Климатологи в общем согласны, что относительно небольшие изменения в содержании СО2 в атмосфере могут вызвать и вызывают существенные изменения климата. Углекислота прозрачна для приходящей энергии видимого солнечного излучения, но, подобно стеклу, поглощает инфракрасное излучение, испускаемое земной поверхностью; возникает, так называемый, “парниковый эффект”, приводящий к увеличению температуры биосферы в целом. Важнейшие предполагаемые последствия повышения температуры - таяние полярных ледяных шапок и установление тропического климата на всей Земле. Подсчитано, что если растает весь антарктический лед, то уровень моря всего за 400 лет поднимается более чем на 120 м. Такое изменение было бы, конечно, катастрофой для человечества - поверхность суши сократится, под водой окажутся низменные территории Европы, прибрежные районы Китая и Индии, запасы пресной воды сократятся на одну треть. Другим следствием повышения температуры может быть увеличившееся испарение с увеличением облачного покрова, что снизит отражательную способность Земли, - это в итоге приведет к еще большему повышению температуры.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
