

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК 637.1:547
СПЕЦИФИЧНОСТЬ ДЕЙСТВИЯ СИАЛОВЫХ КИСЛОТ
В БИОЛОГИЧЕСКИХ СРЕДАХ
Часть 2. ФУНКЦИИ СИАЛОВЫХ КИСЛОТ ПРИ ФОРМИРОВАНИИ МОЛОЧНЫХ СГУСТКОВ
, к. т.н.
ГНУ Сибирский НИИ сыроделия Россельхозакадемии, г. Барнаул
sibniis. *****@***ru
В последнее время особое внимание в странах с развитым сыроделием стало уделяться содержанию κ-казеина в молоке, предназначенном для производства сыра. Еще в 70-х годах прошлого века было установлено, что на качество сыра оказывает влияние не только содержание в молоке казеина, но и соотношение его фракций и генотипов. Так, увеличение доли κ-казеина в молоке на 20 % способствует повышению прочности сычужного сгустка при повышении общего содержания белка на 0,1% [1].
В связи с этим сравнительно новая область генной инженерии – трансгенез – вплотную подошла к решению технологических проблем сыроделия. Наибольший интерес исследователей вызывает генетический ВВ-вариант κ-казеина. Именно с ним связывают перспективы совершенствования производства сыра. Все это и породило в Англии идею создать национальное стадо коров, дающих молоко с этим «чеддерным» вариантом κ-казеина [2,3].
В нашей стране сейчас также считают необходимым типирование коров и быков-производителей по гену κ-казеина путём проведения маркер-направленной селекции при отборе животных, дающих молоко высокого качества [3,4].
Фракция κ-казеина отличается от остальных казеиновых фракций тем, что является гликопротеином, т. е включает в свой состав углеводы и сиаловую кислоту, но содержит мало фосфора.
Основной функцией κ-казеина является стабилизация казеиновой мицеллы. Выполнению этой функции способствует то, что гидрофобные участки к-казеина погружены во внешний слой мицеллы, а гидрофильные экспонированы наружу, создавая электростатический и стерический барьерный слой на поверхности мицеллы. Кроме того, κ-казеин играет роль интерфейса между гидрофобными и Са2+-чувствительными участками α- и β-казеинов и водным окружением казеиновой мицеллы.
Пептидная связь Phe105-Met106 в молекуле κ-казеина с высокой скоростью гидролизуется химозином. Продуктами гидролиза являются пара-κ-казеин (ф.1-105) и гликомакропептид (ф.106-169). Интересно также отметить, что в 8 генетических вариантах κ-казеина из 11 известных, аминокислотные замены локализованы на дистальном участке гликомакропептида (ГМП) и относительно удалены от пептидной связи, гидролизуемой химозином. Эти мутации расположены на отрезке последовательности 136-155 и приходятся на вытянутую часть молекулы, которая выступает в роли физического препятствия коагулирующему воздействию, до момента атаки химозина [цит по 5].
После отщепления ГМП и, следовательно, перехода сиаловых кислот в плазму молока к-казеин теряет функцию «защитного коллоида» и фракция пара-κ-казеина становится чувствительной к ионам кальция. В дальнейшем, в присутствии ионов Са происходит флокуляция казеиновых частиц и молоко свёртывается [6]. Получается, что нечувствительным к ионам кальция является ГМП, несущий большой (–) заряд. Он превращается в белковый компонент плазмы молока.
Сравнивая функции κ-казеина с функциональной специфичностью сиаловых кислот (часть 1), можно сделать логический вывод, что именно сиаловая кислота, входящая в состав гликомакропептида, обуславливает отличительные особенности κ-казеиновой фракции молока.
Если учесть, что плазма молока формируется на основе плазмы крови, то и функциональные особенности сиаловых кислот в молоке имеют тот же механизм действия. Казеиновые частицы и жировые шарики молока представляют собой кинетические отдельности, которые, как и клетки организмов, защищены от внешнего воздействия отрицательным зарядом за счёт локализации на их поверхности сиаловых кислот. При компенсации отрицательного заряда (увеличении концентрации ионов Н+ или катионов) система теряет стабильность и молоко свёртывается.
Например, присутствием сиаловых кислот обусловлена высокая термоустойчивость свежего молока. При рН 6,8-6,6 оно выдерживает кипячение, а свёртывание наступает при температурах выше 130ºС. Именно с этой температуры начинается термическое разложение сиаловой кислоты. Потеря отрицательного заряда на κ-казеине и белках МЖГ приводит к нарушению стабильности системы.
При каком уровне рН свёртывается подкисленное молоко? При сопоставлении этих данных получается довольно интересная картина.
Известно, что при кипячении молока коагуляция казеина наступает при титруемой кислотности порядка 28-30ºТ, что соответствует значениям рН на уровне ~ 6,2-6,1.
По данным [7] изменение физико-химических свойств казеина в первую очередь выражается в изменении степени его дисперсности. В непастеризованном обезжиренном молоке диаметр частиц казеина при естественном сквашивании начинал увеличиваться с 26,5°Т (рН~ 6,15): от частиц с D=67,3 нм до частиц с D=73,1 нм при 34ºТ.
Установлено также [7], что при внесении в молоко культур молочнокислых бактерий в начальной стадии сквашивания величина казеиновых частиц уменьшается. Однако с точки рН 6,14 начинается увеличение их размеров, что означает начало возникновения новых связей между мицеллами казеина. Это приводит к структурным изменениям коллоидной системы казеинового комплекса, направленным в сторону появления новых структурообразований, что в конечном итоге приводит к образованию пространственной гелевой структуры молочного сгустка.
В лаборатории процессов и аппаратов СибНИИС проведены эксперименты по естественному сквашиванию образцов восстановленного обезжиренного молока с массовой долей сухих веществ 30%. Сухое молоко восстанавливали при 37°С и выдерживали в термостате до образования сгустка. Динамика рН и ионного кальция в процессе термостатирования представлена на рис. 1.
Приведённые графические данные констатируют, что система также теряет стабильность на уровне рН 6,17: начинается резкий рост активной кислотности, а с точки рН 6,0 – более интенсивное снижение концентрации ионного кальция, который идёт на построение структурной сетки сгустка.
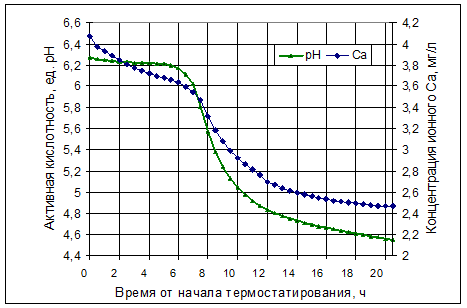
Рисунок 1 Динамика рН и ионного Са в процессе естественного
закисления восстановленного обезжиренного молока
Если выразить полученную динамику в зависимости от активной кислотности, то видно, что происходящие при естественном закислении изменения (рис. 2) имеют ярко выраженные точки перехода скоростей рН и концентрации Са2+.
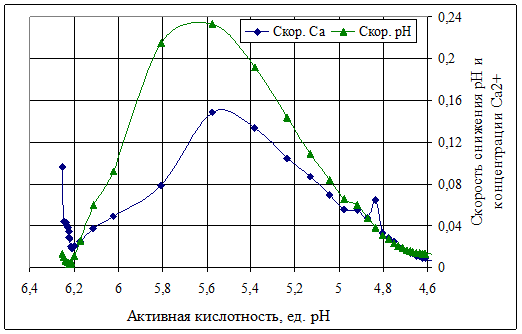
Рисунок 2 Динамика рН и ионного Са в процессе естественного закисления
восстановленного обезжиренного молока в зависимости от уровня активной кислотности
На уровне рН 6,17 происходит затухание одного процесса, а именно потеря стабильности казеинового комплекса, затем следует период формирования структурной сетки (от рН 6,17 до 5,57), далее наблюдается снижение скоростей по мере уплотнения сгустка.
Получается, что в зоне потери стабильности казеинового комплекса (рН 6,2-6,1) приоритетно работает компонент, действия которого не зависят от температуры, ионной силы раствора (массовой доли сухих веществ), вида молока (натуральное или восстановленное) и способа закисления.
Поскольку в этом диапазоне рН по данным [7] наблюдается также минимальный размер казеиновых мицелл, то естественно предположить, что этим компонентом являются сиаловые кислоты, поскольку именно они отвечает за степень проницаемости клеточных мембран и выполняет защитную функцию целостности мицелл. Таким образом, физико-химические свойства и функциональные особенности κ-казеина определяются наличием сиаловых кислот.
Установлено, что в случае естественного сквашивания молока количество свободных сиаловых кислот снижается по мере уменьшения размеров казеиновых частиц и увеличивается с началом процесса агрегации, что можно рассматривать как участие сиаловых кислот в образовании межмолекулярных связей при формировании кислотных сгустков [7].
Ещё одним доказательством участия сиаловых кислот в формировании консистенции молочных продуктов является разница в структурных связях кисломолочных и сычужных сгустков.
Кислотные сгустки характеризуются высокой вязкостью и неразрывностью структуры, что обусловлено присутствием в составе κ-казеина сиаловых кислот, которые в связанном состоянии обуславливают вязкость биологических водных сред. Считается, что кислотные сгустки отличаются от сычужных отсутствием связей через коллоидный фосфат кальция (КФК) [8]. Но тогда напрашивается вопрос: а какие связи присутствуют? Если учесть, что в кисломолочных сгустках массовая доля воды составляет обычно более 80%, то, по всей вероятности, в них преобладают водородные связи. Специфичность действия сиаловых кислот и их функциональные возможности соединяться с белками, жирами и углеводами, по-видимому, тоже вносит существенный вклад в формирование структуры кисломолочных сгустков. Следует учитывать и тот факт, что сиаловые кислоты присутствуют в клетках заквасочных микроорганизмов и после их лизиса также могут стать элементами структурной сетки.
Сычужные сгустки, в которых κ-казеин лишился углеводной части и все казеиновые фракции, включая пара-κ-казеин, характеризуются сродством к Са2+, формируются посредством ионных связей через кальций, что способствует их разрывности и склонности к синерезису.
Что известно об участии сиаловых кислот в формировании структуры сычужных сгустков? С момента разделения к-казеина на углеводсодержащую фракцию (ГМП) и основной компонент, получивший название пара-κ-казеина, начинается процесс образования сычужного сгустка. Считается, что после разрезки сгустка ГМП с сиаловой кислотой уходит в сыворотку и его роль в сыроделии на этом заканчивается.
Исследования по изучению роли сиаловых кислот в процессе сычужного свёртывания молока в бывшем СССР проводились в Литовском филиале ВНИИМС под руководством д. т.н., профессора [7]. Эксперименты проводились на свежем обезжиренном молоке, пастеризованном при 72ºС с выдержкой 15 с.
Результаты проведённой работы позволили констатировать следующее.
1) Количество свободных сиаловых кислот (ССК) в сычужных сгустках из свежего и зрелого молока одной партии выравнивается по мере формирования их структуры, несмотря не некоторые отличия в скорости накопления ССК в процессе сычужного свёртывания (рис. 3).
2) Количество ССК в сформированных сычужных сгустках одинаково вне зависимости от температуры свёртывания (рис. 4). От температуры зависит скорость накопления свободных сиаловых кислот и, соответственно, время формирования сычужного сгустка.
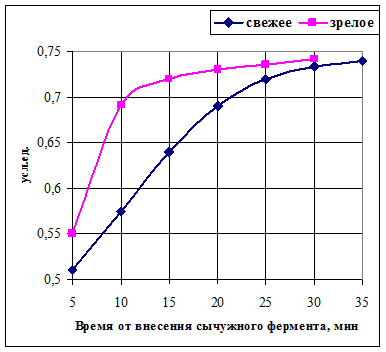
Рисунок 3 Влияние созревания молока на изменение количества свободных сиаловых
кислот во время сычужного свёртывания [ ]
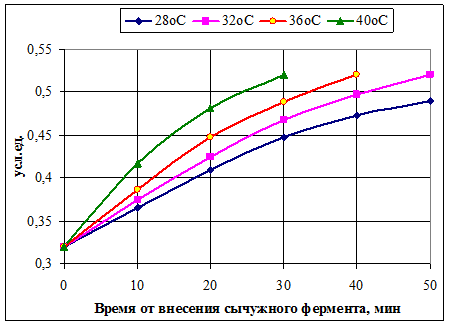
Рисунок 4 Изменение количества свободных сиаловых кислот в зависимости
от температуры сычужного свёртывания молока [7]
3) В наибольшей степени на динамику количества свободных сиаловых кислот при сычужном свёртывании влияет предварительная обработка молока, а именно температура пастеризации (рис. 5)
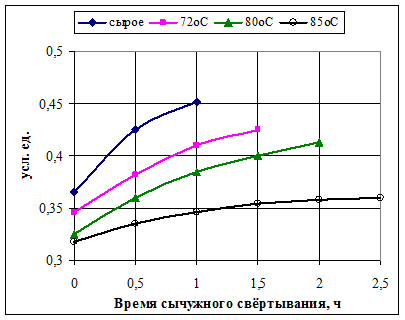
Рисунок 5 Влияние температуры пастеризации молока на изменение количества
свободных сиаловых кислот во время сычужного свёртывания [7]
Таким образом, процесс накопления ССК в большей степени зависит от предистории подготовки молока к свёртыванию, чем от параметров сычужного свёртывания. С повышением температуры пастеризации изменяется состав плазмы молока и стехиометрические параметры системы. Кроме того, молоко лишается ферментативной поддержки [7].
Молоко теряет способность к сычужному свёртыванию, если температура его пастеризации превышает 72ºС. Это связано с тем, что денатурированные сывороточные белки, взаимодействуя с фракцией κ-казеина, закрывают доступ к нему сычужного фермента. Как это сказывается на дисперсности казеиновых частиц, наглядно иллюстрирует рис. 6.
Как и при кислотном свёртывании, диаметр казеиновых частиц сначала уменьшается, а затем увеличивается по мере формирования пространственной структуры сгустка. Однако, чем выше температура пастеризации, тем крупнее исходные (перед свёртыванием) казеиновые частицы и тем дольше по времени протекает процесс формирования сгустка. Это напрямую связано со способностью сиаловых кислот взаимодействовать с самой объёмной фракцией сывороточных белков – лактоглобулином, тем самым предотвращая доступ сычужного фермента к κ-казеину.
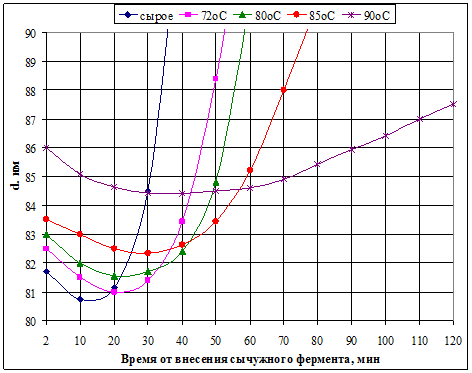
Рисунок 6 Изменение дисперсности казеинового комплекса при сычужном свёртывании
молока, пастеризованного при разных температурах [7]
Признаком превращения молока из золя в гель является формирование в среде длинных и тонких молекул [9]. Именно такими и являются молекулы казеинов. Согласно реоморфной теории (Holt a. e., 1992) они собраны в мицеллярные образования для выполнения физиологической функции – быстрого изолирования кальций-фосфатных нанокластеров в выделительных везикулах молочных желез млекопитающих. Перенос кальция в форме мицелл казеина является для молочных желез наименее безопасным способом его транспорта [10].
Образование сгустка – это расформирование мицелл казеина и преобразование белков в нитеобразную форму. В настоящее время известно несколько десятков моделей структуры казеиновой мицеллы. Естественно, что процесс формирования сгустков до сих пор остаётся дискуссионным. Например, можно привести следующую трактовку формирования сычужного сгустка.
«В начале агрегации мицеллы казеина чаще всего образуют цепочки, в которых они являются звеньями, соединенными с соседними в нескольких точках поверхности. Затем площадь контактов увеличивается и цепочки постепенно переходят в пряди шириной в 1-4 и длиной до 10 мицелл.
В отличие от цепочек, ширина прядей по всей длине выравнивается за счет расширения участков соединения мицелл. К началу образования видимого невооруженным взглядом сгустка пряди связываются друг с другом поперечными связями с образованием мелкоячеистой негомогенной структуры с диаметром ячеек до 10 мкм и с порами размером около 0,5 мкм. Стенки ячеек образованы агрегированными параказеиновыми мицеллами, имеющими спиралеобразную форму. В местах соединения связей образуются узлы. Внутри ячеек содержатся жировые шарики и сыворотка.
Далее площадь контакта мицелл друг с другом увеличивается; они образуют конгломераты, между которыми образуются поры до 10 мкм в диаметре, а затем конгломераты постепенно сливаются в сплошную массу параказеина. Весь сгусток как бы становится гигантской параказеиновой «мицеллой» с многочисленными порами» [цит. по 8].
Установлено, что на начальном этапе сычужного свёртывания происходит уменьшение размеров казеиновых частиц (рис.6) и, соответственно, снижение вязкости молока. Повышение дисперсности казеиновых частиц объясняется ферментативным гидролизом κ-казеина: отщеплением ГМП и снятием гидратных оболочек с мицелл.
Согласно [7] во время формирования сычужного сгустка содержание ССК увеличивается независимо от направленности изменения дисперсности казеиновых частиц. При этом к моменту образования сгустка процесс их накопления заканчивается. В этой связи количество ССК в определённой степени характеризует изменение стабильности казеинового комплекса во время отдельных операций. С учётом этого КССК на разных этапах сычужного свёртывания можно принимать в качестве показателя изменения структуры казеинаткальцийфосфатного комплекса.
установлено, что между временем образования сычужного сгустка и количеством ССК существует функциональная зависимость:
У = 8,25 – 15,8х,
где У – время образования сгустка; х – количество ССК.
Учитывая вышесказанное можно сделать вывод, что при формировании сычужных сгустков сычужный фермент выполняет функцию снижения отрицательной зарядности мицелл, что приводит к снижению ИЭТ параказеина ~ на 0,6 ед. рН (∆ рН).
Если от исходной активной кислотности молока отнять ∆ рН = 0,6 ед., то мы получим: 6,7 – 0,6 = 6,1. Именно в этой области рН начинается процесс увеличения количества ССК в кислотных сгустках. Действие сычужного фермента выражается в том, что осуществляя «скачок по рН» (∆= 0,6 ед.), он ускоряет процесс накопления ССК и далее механизм их действия идёт по одному сценарию в сычужных и кислотных сгустках.
До рН 6,14 [7] при кислотном свёртывании наблюдается уменьшение диаметра казеиновых мицелл, снижение вязкости и уменьшение количества ССК. Данную область рН (6,2-6,1) называют также сферой действия белкового буфера в молоке. По аналогии с действием фосфатного буфера можно предположить, что если ионы Н+ могут связываться в соответствии с реакцией: СаНРО4 + Н+ → Са(Н2РО4)2, то при работе белкового буфера свободные сиаловые кислоты могут расходоваться на поддержание отрицательного заряда на поверхности казеиновых мицелл. Здесь происходит процесс, называемый буферностью по белку, т. е. ССК усиливают защитную оболочку мицелл.
Уменьшение размеров казеиновых мицелл до рН 6,14 соответствует тому факту, что при снижении рН наблюдается процесс прилегания «волоскового» слоя к поверхности мицелл [11]. А «волосковый слой», выступающий с поверхности мицелл, как известно, представляет собой гликомакропептидную часть k-казеина [12].
При достижении рН уровня 6,2-6,1 буферная ёмкость исчерпывается и происходит процесс отщепления сиаловых кислот от κ-казеина, что экспериментально фиксируется как повышение количества ССК [7] и повышение скорости снижения рН (рис. 1).
Исходя из вышеизложенного можно сделать вывод, что регулятором процесса расформирования мицелл являются сиаловые кислоты, а увеличение размеров мицелл в процессе формирования сгустка можно трактовать как процесс ослабления внутримицеллярных связей, способствующих переходу системы на новый уровень структурообразования.
Участие сиаловых кислот в формировании кисломолочных сгустков, учитывая тот факт, что их структурообразование происходит с участием всех компонентов молока, а сиаловые кислоты входят в состав многих из них, можно считать неоспоримым.
Что касается роли сиаловых кислот в процессе сычужного свёртывания с последующим разделением системы на сгусток и сыворотку, то этот вопрос остаётся практически открытым и требует дальнейших исследований. Вполне возможно, что выявление механизма специфичности действия сиаловых кислот, в частности, особенностей их взаимодействия с сывороточными белками, позволит снизить степень перехода сухих веществ в сыворотку и повысить выход продукции сыроделия.
Можно с уверенностью предположить, что специфичность действия сиаловых кислот, входящих в состав κ-казеина и белков мембран жировых глобул, оказывает значительное влияние на процессы биотрансформации компонентов молока при его переработке, а, следовательно на качество и выход молочной продукции.
Список литературы
1 Руководство по молочному делу и гигиене молока.- М.: Россельхозиздат, 1980.– 205 с.
2 http://www. *****/archive/articles/9572/ Стадо для чеддера / Наука и жизнь / Архив журнала «Наука и жизнь» / Наука на марше / Интервью № 8, 1999 год
3http://www. *****/phorum/viewtopic. html&f=6&t=2121&archive=1&search=%F1%FB%F0&page=2
4 http://www. yariks. info/arhive. php? c=44&pc=71&m=202
5 , Кригер и биохимические свойства казеинов коровьего молока. 4. Каппа-казеины. // Сыроделие и маслоделие, 2008. – № 5. – С. 53-56.
6 , Горбатова молока и молочных продуктов /под ред. проф. . – Л., Изд-во Ленингр. ун-та, 1974. – 260 с.
7 Раманаускас главы физической химии сыроделия / Монография. Каунас, 2004. – 142 с.
8 Гудков : технологические, биологические и физико-химические аспекты / под ред. – М.: ДеЛи принт, 2003. – 800 с.
9 Asimov I. The genetic code. – London, John Murray, 1964. – 161 s.
10 Holt C. Structure and stability of bovine casein micelles.// Adv. Prot. Chem. – 1992, V 43 – p. 63-151.
11 Horne D. S., Parker T. G., Dalgleish D. G. Casein micelles, polycondensation and fractals.// Food Colloids, Spec. Publ. No. 75. R. Soc. Chem., London, 1989 – p. 400-405.
12 Dalgleish D. G. A mechanism for the chymosin-induced flocculation of casein micelles// Biophysical Chemistry. 1980, v. 11, p. 147-155.
СПЕЦИФИЧНОСТЬ ДЕЙСТВИЯ СИАЛОВЫХ КИСЛОТ
В БИОЛОГИЧЕСКИХ СРЕДАХ
Часть 2. ФУНКЦИИ СИАЛОВЫХ КИСЛОТ ПРИ ФОРМИРОВАНИИ МОЛОЧНЫХ СГУСТКОВ
, к. т.н.
ГНУ Сибирский НИИ сыроделия Россельхозакадемии, г. Барнаул
sibniis. *****@***ru
Рассмотрены функциональные особенности κ-казеина относительно стабилизации казеиновых мицелл, дана сравнительная оценка функциональных особенностей сиаловых кислот при кислотном и сычужном свёртывании молока. Показано, что количество свободных сиаловых кислот в определённой степени характеризует изменение стабильности казеинового комплекса во время отдельных операций.
