
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
пробы должны обрабатываться вместе (одновременно), чтобы условия проведения реакции были одинаковыми. Расчет ведут по отношению к стандарту. При этом результат тем точнее, чем ближе концентрации опытных и стандартных проб.
№14.Прицип метода диализа.
Диализ - искусственное очищение крови и других жидкостей человеческого тела от скопившихся шлаков, когда органы самого человека неспособны нормально выполнять эти функции. Обычно кровь фильтруется в почках, которые задерживают полезные вещества, а вредные выделяют с мочой. При заболеваниях почек, когда одна или обе почки не функционируют, больному необходим диализ. Существуют два метода диализа. Первый - очищение крови через брюшину (перитониальный) в самом теле, второй - подключение к искусственной почке.
Принцип действия диализа
Принцип действия диализа основан на законах осмоса. Для того, чтобы эти законы действовали, необходимы очень тоненькие мембраны, проницаемые только для воды и определенных веществ, растворенных в ней. Когда с обеих сторон этих мембран образуется различная концентрация жидкости, тогда эти жидкости пытаются проникнуть сквозь мембрану, чтобы выровнять уровень концентрации и установить равновесие: если с одной стороны мембраны каких-либо веществ больше, чем с другой, часть их попадает на другую сторону. Естественные мембраны человеческого тела - это легкие, брюшина и капилляры почек; эти мембраны пропускают определенные вещества, а другие составные части крови в ней и остаются. Точно таков механизм действия диализа, который назначается при нарушении функции почек или отравлениях (грибами, лекарствами). Существует много людей, которым диализ повторяют несколько раз в неделю. При тяжелых заболеваниях почек гемодиализ (искусственная почка) еще может обеспечить почти нормальную жизнь, однако известно, что лучшее решение - это пересадка почки (трансплантация).
№15.Классификация белков.
В настоящее время еще не разработана стройная система номенклатуры и классификации белков. Традиционная классификация белков по группам, основанная, скорее, на случайных показателях (физико-химические свойства, форма молекул, локализация и происхождение, аминокислотный состав), уже не отвечает полностью возросшему уровню знаний о их структуре и функциях. Из огромного количества природных белков структура и функции расшифрованы для относительно небольшого числа (не более нескольких сотен), и поэтому структура и функции белков пока не могут служить основой для их рациональной классификации. Пожалуй, только для одной группы белков, обладающих способностью катализировать химические реакции, т. е. ферментов, разработана стройная система номенклатуры и классификации, в основу которой положены типы катализируемых химических реакций и химическая природа реагирующих веществ. Однако полностью идентифицированные до сих пор ферменты также составляют незначительную долю белков (ферментов описано более 3000). Тем не менее функциональный принцип, рекомендуемый некоторыми авторами, хотя и не может служить универсальной основой для классификации всех белков, представляет определенный интерес. В соответствии с функциональным принципом различают 12 главных классов белков: 1) каталитически активные белки (ферменты); 2) белки-гормоны (хотя есть и стероидные гормоны); 3) белки-регуляторы активности генома; 4) защитные белки (антитела, белки свертывающей и антисвертывающей систем крови); 5) токсические белки; 6) транспортные белки; 7) мембранные белки; 8) сократительные белки; 9) рецепторные белки; 10) белки-ингибиторы ферментов; 11) белки вирусной оболочки; 12) белки с иными функциями.
Были предприняты также попытки классифицировать белки, исходя из особенностей вторичной и третичной структуры. В соответствии с этим различают α-, β-, α+β- и α/β-белки. α-Белки содержат только α-спирали (не менее 60%), β-белки – только β-структуры (не менее двух антипараллельных цепей), α+β-белки – те и другие структуры в пределах одной полипептидной цепи (пример – молекулы лизоцима), а класс α/β-белков содержит множество α- и β-структур, чередующихся вдоль полипептидной цепи или домена (см. рис. 1.19). Домены создаются объединением и чередованием α-спиралей и β-слоев, между которыми открываются более рыхлые структуры.
Учитывая необходимость для студента-медика основательного знакомства с отдельными группами белков, которые могут служить предметом изучения в его будущей профессиональной деятельности (например, белки крови), приводим старую классификацию белков с краткой характеристикой новых данных о структуре, составе и свойствах отдельных представителей. Согласно этой классификации, обширный класс белковых веществ в зависимости от химического состава делят на простые и сложные белки.
Простые белки построены из остатков аминокислот и при гидролизе распадаются соответственно только на свободные аминокислоты.
Сложные белки – это двухкомпонентные белки, которые состоят из какого-либо простого белка и небелкового компонента, называемого про-стетической группой. При гидролизе сложных белков, помимо свободных аминокислот, освобождается небелковая часть или продукты ее распада.
Простые белки в свою очередь делятся на основании некоторых условно выбранных критериев на ряд подгрупп: протамины, гистоны, альбумины, глобулины, проламины, глютелины и др. Классификация сложных белков (см. главу 2) основана на химической природе входящего в их состав небелкового компонента. В соответствии с этим различают фосфопротеины (содержат фосфорную кислоту), хромопротеины (в состав их входят пигменты), нуклеопротеины (содержат нуклеиновые кислоты), гликопротеины (содержат углеводы), липопротеины (содержат липиды) и металлопротеины (содержат металлы).
№16.Характеристика сложных белков.
Простые белки построены только из аминокислот. Сложные белки построены из двух компонентов - простой белок и небелковое вещество, называемое простетической группой. Простетические группы прочно связаны с белковой частью молекулы.
Классификация сложных белков зависит от строения простетической группы.
Гликопротеины (содержат углеводы).
Липопротеины (содержат липиды).
Фосфопротеины (содержат фосфорную кислоту).
Хромопротеины (содержат окрашенную простетическую группу).
Металлопротеины (содержат ионы различных металлов).
Нуклеопротеины (содержат нуклеиновые кислоты).
№17.Нуклеопротеины.
Нуклеопротеины состоят из белков и нуклеиновых кислот. Последние рассматриваются как простетические группы. В природе обнаружено 2 типа нуклеопротеинов, отличающихся друг от друга по составу, размерам и физико-химическим свойствам,– дезоксирибонуклеопротеины (ДНП) и рибонуклеопротеины (РНП). Названия нуклеопротеинов отражают только природу углеводного компонента (пентозы), входящего в состав нуклеиновых кислот. У РНП углевод представлен рибозой, у ДНП – дезоксирибозой. Термин «нуклеопротеины» связан с названием ядра клетки, однако ДНП и РНП содержатся и в других субклеточных структурах. Следовательно, речь идет о химически индивидуальном классе органических веществ, имеющих своеобразные состав, структуру и функции независимо от локализации в клетке. Доказано, что ДНП преимущественно локализованы в ядре, а РНП – в цитоплазме. В то же время ДНП открыты в митохондриях, а в ядрах и ядрышках обнаружены также высокомолекулярные РНП.
Пристальное внимание исследователей привлечено к структуре и функции макромолекул, включающих комплексы белков и нуклеиновых кислот. Этот особый интерес вызван тем, что многообразие проявлений жизни непосредственно связано с этими полимерными молекулами. Биохимики имеют достаточно оснований для утверждения, что природа синтезированных в клетках белков зависит в первую очередь от природы ДНП, точнее ДНК, а свойства живых организмов, как и структурная организация субклеточных органелл, клеток и целостного организма, определяются свойствами синтезированных белков.
ДНК хранит наследственную информацию. Подтверждением этого служит явление трансформации, наблюдаемое у бактерий и открытое также в культуре клеток человека. Сущность явления заключается в превращении одного генетического типа клеток в другой путем изменения природы ДНК. Так, удалось получить штамм капсулированных и вирулентных пневмококков из исходного штамма, не обладающего этими признаками, путем внесения в среду ДНК, выделенной из капсулированного (и вирулентного) штамма. С нуклеопротеинами и соответственно нуклеиновыми кислотами непосредственно связаны, кроме того, такие биологические процессы, как митоз, мейоз, эмбриональный и злокачественный рост и др.
У большинства клеток эукариот, когда ядро находится в интерфазе, из ДНК и белковых молекул образуются так называемые филаменты – нити, имеющие меняющуюся толщину (в среднем около 10 нм, реже 2 нм). Оказывается, что толщина филаментов определяется наличием или отсутствием белков, окружающих двухспиральную структуру ДНК, а длина их – молекулярной массой ДНК. Известно, что одна хромосома содержит одну молекулу ДНК, имеющую длину несколько сантиметров. Вообще ДНП входит в состав мононуклеосом, являющихся составной частью хромосомы. Таким образом, в состав хроматина входят молекула ДНК, пять различных классов белков – гистонов и так называемые негистоновые белки. Количество ДНК в ядре составляет до 6 пг (10–12 г) на одну клетку у животных. У E. coli содержание ДНК равно 0,01 пг.
Относительно белкового состава ДНП известно, что все 5 классов гистонов различаются по размерам, аминокислотному составу и величине заряда (всегда положительный). Так, выделяют гистоны, богатые лизином (H1), молекулярная масса которых составляет в среднем 20000, и богатые аргинином с мол. массой до 15000. Они обозначаются следующими символами:
H1 – богатые лизином,
Н2А – богатые аргинином и лизином,
Н2В – умеренно богатые аргинином и лизином,
Н3 – богатые аргинином,
Н4 – богатые глицином и аргинином.
Природа негистоновых белков пока не достаточно выяснена. Негисто-новые белки в настоящее время интенсивно изучаются. В их состав входят сложные белки, ферменты, а также регуляторные белки. По своим свойствам последние отличаются от гистонов и представлены кислыми белками.
В различных нуклеопротеинах количество нуклеиновой кислоты колеблется от 40 до 65% (например, в рибосомах про - и эукариот). В вирусных нуклеопротеинах количество нуклеиновых кислот не превышает 2–5% от общей массы. Так, у вируса табачной мозаики (ВТМ) на долю РНК, правда, с огромной молекулярной массой – около 2 приходится всего около 2%. Остальная часть этой гигантской вирусной частицы приходится на долю однотипных белковых субъединиц (рис. 2.3). Ионная связь между РНК и белковыми молекулами ВТМ весьма непрочная и легко разрывается даже в «мягких» условиях, что позволяет отделить РНК от белка. Интересно, что после удаления разрывающего ионную связь агента при смешивании этих продуктов происходят полная регенерация исходного ВТМ, восстановление всех его физических параметров и биологических свойств, включая способность поражать зеленый лист. Это явление самосборки, впервые открытое у ВТМ, в дальнейшем было обнаружено также у бактериофагов, представленных нуклеопротеинами. Акад. и одновременно М. Номура разделили 70S рибосомы (рибонуклеопротеины) на их составляющие и разработали условия для самосборки полноценных функционирующих рибосом. В основе этого удивительного явления самосборки лежит, по-видимому, программа, содержащаяся в первичной структуре как белка, так и нуклеиновой кислоты и определяющая, какое количество белковых молекул и в какой последовательности должно присоединиться к единственной молекуле РНК (в случае ВТМ) или к 3 молекулам РНК (в рибосомах), чтобы обеспечить высокую точность реконструкции надмолекулярных структур.
В настоящее время и ядерный хроматин (ДНП), и рибосомы, и вирусные нуклеопротеиды обычно рассматривают именно как надмолекулярные комплексы или структуры, а отнесение этих образований в раздел «Сложные белки» – в значительной степени дань традиции.
№18,19.Гликопротеины.
Гликопротеины – сложные белки, содержащие, помимо простого белка или пептида, группу гетероолигосахаридов. В настоящее время их принято называть гликоконъюгатами. В состав гликоконъюгата входит углеводный компонент (гликановая фракция), ковалентно связанный с неуглеводной частью (агликановая фракция), представленной белком, пептидом, аминокислотой или липидом.
Повышенный интерес к науке об углеводах – гликобиологии – в настоящее время объясняется открытием существенной роли изменений структуры гликоконъюгатов в развитии таких болезней, как рак, иммунодефицит человека, ревматоидные артриты, астма и др. Оказалось, что нарушение реакции гликозилирования (см. главу 14) двух главных классов глико-конъюгатов (гликопротеинов и ганглиозидов) приводит или к накоплению предшественников этих веществ, или к синтезу «укороченных» сахарных цепей гликоконъюгатов. Более того, установлено, что во взаимодействии между некоторыми вирусами и клетками-мишенями главную роль играют углеводные компоненты. В частности, гликопротеин gp120 вируса иммунодефицита человека (содержит большой процент углевода) имеет высокое сродство к гликопротеину CD4Т-лимфоцита. В этом взаимодействии, узнавании, являющемся высокоспецифичным, гликозилированные фрагменты, вероятнее всего, играют важную патогенетическую роль. Известно также, что при ревматоидных артритах часто синтезируются аномальные антитела (аномальные иммуноглобулины – все гликопротеины) с необычайно короткими сахарными цепями, что вызывает стимуляцию иммунной системы против самого организма. Из этих примеров видно, что, помимо гликобиологии, наступило время признания и таких наук, как гликопа-тология и гликотерапия.
Помимо гликопротеинов, различают также протеогликаны, состоящие из белка и гликозаминогликанов (прежнее название мукополиса-хариды); последние состоят из цепей сложных углеводов: аминосахаров, уроновых кислот, серной кислоты и отдельных моносахаридов.
К типичным гликопротеинам относят большинство белковых гормонов, секретируемые в жидкие среды организма вещества, мембранные сложные белки, все антитела (иммуноглобулины), белки плазмы крови, молока, овальбумин, интерфероны, факторы комплемента, группы крови, рецеп-торные белки и др. Из этого далеко не полного перечня гликопротеинов видно, что все они выполняют специфические функции: обеспечивают клеточную адгезию, молекулярное и клеточное узнавание, антигенную активность опухолевых клеток, оказывают защитное и гормональное, а также антивирусное действие.
Химический состав гликопротеинов более или менее установлен, структура определена только у ряда из них. К полипептиду присоединяются гетероолигосахаридные цепи, содержащие от 2 до 10, реже 15 мономерных остатков гексоз (галактоза и манноза, реже глюкоза), пентоз (ксилоза, арабиноза) и конечный углевод, чаще всего представленный N-ацетилга-лактозамином, L-фукозой или сиаловой кислотой; в отличие от про-теогликанов гликопротеины не содержат уроновых кислот и серной кислоты.
Типы связей между углеводными компонентами и белками определены только у ряда гликопротеинов, аминокислотный состав и структура которых известны (иммуноглобулины, гормоны); они включают О-гликозидные связи (с ОН-группами серина, треонина и оксилизина), N-гликозидные связи (с амидными группами аспарагина, реже глутамина или ω-NH2-группами лизина и аргинина) и эфирные гликозидные связи со свободными СООН-группами глутаминовой и аспарагиновой кислот.
Синтез гликопротеинов осуществляется в рибосомах эндоплазматиче-ского ретикулума (в цистронах), затем присоединяются сахарные цепи (постсинтетическое гликозилирование), и далее белок транспортируется до биомембран клетки и включается в состав мембранных белков или секре-тируется.
Углеводные компоненты соединены ковалентно с азотом аспарагина молекулы белка. Однако предварительно олигосахаридная часть соединяется с липидным переносчиком – долихолфосфатом (липид, содержащий от 15 до 20 изопреновых остатков) и переносится на полипептидную цепь в эндоплазматическом ретикулуме, при этом транспортер освобождается:
Долихолфосфат (n = 15-30)
Синтезированные гликопротеины далее переносятся в аппарат Гольджи, где осуществляются окончательное гликозилирование и сортировка по назначению.
Структура одного из нескольких гетероолигосахаридных остатков в молекуле гликопротеинов, в частности иммуноглобулинов, может быть представлена в виде следующей схемы (использованы сокращения: Глю – глюкоза, NАцГлюА – N-ацетилглюкозамин; Гал – галактоза; Ман – манно-за; NАцНейр – N-ацетилнейраминовая кислота):
Интерфероны. Интерфероны – это ингибиторы размножения многих типов вирусов. Открыто несколько типов интерферонов (α, β и γ), некоторые из них получены методами генетической инженерии. Это сравнительно небольшие сложные белки с мол. массой у разных видов животных и человека от 25000 до 38000–40000). Они образуются в клетке в ответ на внедрение вирусной нуклеиновой кислоты, ограничивая вирусную агрессию (инфекцию). Известно также, что группа видоспецифических α-интерфе-ронов синтезируется макрофагами, в то время как γ-интерферон продуцируется Т-клетками и стимулируется интерлейкином-2 (см. «Лимфо-кины»). Показано также, что γ-интерферон в свою очередь повышает цитотоксическую активность макрофагов, Т-клеток и естественных клеток-киллеров. Интерфероны наделены антипролиферативной активностью и считаются основными защитными белками не только против вирусной инфекции, но и при опухолевых поражениях.
Следует отметить, однако, что до сих пор не раскрыты молекулярные механизмы, при помощи которых интерфероны тормозят размножение вирусов. Известно только, что интерфероны ингибируют биосинтез всех белков (и хозяйских, и вирусных), вероятнее всего, на уровне процесса трансляции. Возможно, что интерферон индуцирует синтез особого белка-ингибитора, который затем связывается с рибосомами и блокирует трансляцию, или интерферон переводит один из активных эукариотических белковых факторов инициации в неактивный фактор путем фосфорилиро-вания.
Иммуноглобулины. Иммуноглобулины, или антитела, также относятся к классу гликопротеинов, выполняют защитную функцию, обезвреживая поступающие в организм чужеродные вещества – антигены любой химической природы. Синтезируются иммуноглобулины плазматическими клетками, образовавшимися из лимфоцитов. Учение об иммунитете оформилось в самостоятельную науку – иммунологию, изучающую структуру и функции антител вообще и иммуноглобулинов в частности. Мы представим современные сведения о некоторых физико-химических свойствах и структуре иммуноглобулинов человека (табл. 2.4). Различают 5 классов иммуноглобулинов: IgG, IgM, IgA, IgD и IgE. Детально изучены структура и функция IgG.
Разные классы иммуноглобулинов сильно различаются не только по молекулярной массе, но и по концентрации в крови; имеются данные, что различаются они и по биологическим свойствам.
Подробно изучена структура IgG. Он имеет Y-образную форму и тет-рамерное строение; состоит из двух идентичных легких L-цепей (от англ. light) и двух идентичных тяжелых Н-цепей (от англ. heavy) с мол. массой 23000–24000 и 50000–70000 соответственно. Известно также, что каждая из этих цепей имеет 2 типа доменов – вариабельные (V) участки, состоящие из 108 аминокислотных остатков, и константные (С) участки, состоящие из 110 и 350 аминокислотных остатков соответственно в L - и Н-цепях (рис. 2.5).
Из других гликопротеинов, выполняющих ряд важнейших биологических функций, следует отметить все белки плазмы крови (за исключением альбуминов), трансферрин, церулоплазмин, гонадотропный и фолликуло-стимулирующие гормоны, некоторые ферменты, а также гликопротеины в составе слюны (муцин), хрящевой и костной тканей и яичного белка (овомукоид). Углеводные компоненты, помимо информативной функции, значительно повышают стабильность молекул, в состав которых они входят, к различного рода химическим, физическим воздействиям и предохраняют их от действия протеиназ, определяя тем самым биологическую роль гликопротеинов. Являясь составной частью клеточной мембраны, гликопротеины участвуют, кроме того, в иммунологических реакциях, ионном обмене, процессах межклеточной адгезии и т. д.
№20.Протеогликаны.
Протеогликаны — гликопротеины с высокой степенью гликозилирования, углеводные остатки которых представляют собой длинные неразветвленные полисахаридные цепи — гликозаминогликаны, образованные чередующимися остатками гексозамина и уроновой кислоты (глюкуроновой, идуроновой или галактуроновой) либо галактозы. Гликозаминогликановые цепи гликопротеинов зачастую сульфированы[1].
Протеогликаны являются одним из основных компонентов внеклеточного матрикса соединительной ткани.
Протеогликаны состоят из сердцевинных белков (лат. core protein) с молекулярной массой от 10 до 600 кДа, с которыми ковалентно связаны несколько гликозаминогликановых цепей (до 80 гликозидных звеньев массой докДа); число связанных с белком полисахаридных цепей варьирует от одной (декорин фибробластов) до ста и выше (аггрекан хрящевой ткани). Такое строение обуславливает высокую молекулярную массу протеогликанов (у аггрекана до 5 МДа), при этом на белковую часть приходится ~5-10% и на углеводную ~90-95% массы.
Сердцевинные белки протеогликанов, как и другие белки, синтезируются рибосомами шероховатого эндоплазматического ретикулума и транспортируются в аппарат Гольджи, где происходит их гликозилирование. На первой стадии происходит «наращивание» на сериновом остатке сердцевинного белка «связующего тетрасахарида» (англ. proteoglycan linkage tetrasaccharide) «ксилоза-галактоза-галактоза-глюкуроновая кислота»(ß1-4Xyl ß1-3Gal ß1-3Gal GlcA), после чего соответствующими гликозилтрансферазами осуществяется наращивание специфичной для данного протеогликана гликозиламиногликановой цепи.
№21.Мукополисахаридозы.
Мукополисахаридозы или сокращенно МПС, или MPS (от (мукополисахариды + -ōsis)) — группа метаболических заболеваний соединительной ткани, связанных с нарушением обмена кислых гликозаминогликанов (GAG, мукополисахаридов), вызванных недостаточностью лизосомных ферментов обмена гликозаминогликанов. Заболевания связаны с наследственными аномалиями обмена, проявляются в виде «болезни накопления» и приводят к различным дефектам костной, хрящевой, соединительной тканей.
Виды заболеваний
В зависимости от характера ферментативного дефекта выделяют несколько основных типов мукополисахаридозов:
I тип — синдром Гурлер (мукомполисахаридоз I H — Hurler), синдром Гурлера-Шейе (мукополисахаридоз I H/S — Hurler-Scheie), синдром Шейе (мукополисахаридоз I S — Scheie). Обусловлен дефицитом альфа-L-идуронидазы (фермент катаболизма мукополисахаридов). Заболевание постепенно приводит к накоплению в тканях гепарансульфата и дерматансульфата. Выделяют три фенотипа: синдром Гурлера, синдром Шейе и синдром Гурлера-Шейе.
II тип — синдром Хантера
III тип — синдром Санфилиппо
IV тип — синдром Моркио
V тип — синдром Шейе
VI тип — синдром Марото—Лами
VII тип — синдром Слая дефицит р-глюкуронидазы
№22.Хромопротеины.
Сложные белки, как было отмечено, содержат два компонента – простой белок и небелковое вещество. Последнее называют простетической группой (от греч. prostheto – присоединяю). Простетические группы, как правило, прочно связаны с белковой молекулой. Далее представлены сведения о химической природе и биологической роли некоторых сложных белков.
ХРОМОПРОТЕИНЫ
Хромопротеины (от греч. chroma – краска) состоят из простого белка и связанного с ним окрашенного небелкового компонента. Различают гемопротеины (содержат в качестве простетической группы железо), маг-нийпорфирины и флавопротеины (содержат производные изоаллоксазина). Хромопротеины наделены рядом уникальных биологических функций: они участвуют в таких фундаментальных процессах жизнедеятельности, как фотосинтез, дыхание клеток и целостного организма, транспорт кислорода и диоксида углерода, окислительно-восстановительные реакции, свето-и цветовосприятие и др.
Таким образом, хромопротеины играют исключительно важную роль в процессах жизнедеятельности. Например, подавление дыхательной функции гемоглобина путем введения оксида углерода (СО) либо утилизации (потребление) кислорода в тканях путем введения синильной кислоты или ее солей (цианидов), ингибирующих ферментные системы клеточного дыхания, моментально приводит к смерти организма.
Хромопротеины являются непременными и активными участниками аккумулирования энергии, начиная от фиксации солнечной энергии в зеленых растениях и утилизации ее до превращений в организме животных и человека. Хлорофилл (магнийпорфирин) вместе с белком обеспечивает фотосинтетическую активность растений, катализируя расщепление молекулы воды на водород и кислород (поглощением солнечной энергии). Гемопротеины (железопорфирины), напротив, катализируют обратную реакцию – образование молекулы воды, связанное с освобождением энергии.
№23.Строение гемоглобина.
Гемоглобин-белок, включающий 4 гемсодержащие белковые субъединицы. Между собой протомеры соединяются гидрофобными, ионными, водородными связями по принципу комплементарности. При этом они взаимодействуют не произвольно, а определенным участком - контактной поверхностью. Этот процесс высокоспецифичен, контакт происходит одновременно в десятках точек по принципу комплементарности. Взаимодействие осуществляют разноименно заряженные группы, гидрофобные участки, неровности на поверхности белка.
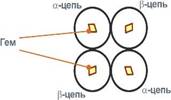
Белковые субъединицы в нормальном гемоглобине могут быть представлены различными типами полипептидных цепей: α, β, γ, δ, ε, ξ (соответственно, греч. - альфа, бета, гамма, дельта, эпсилон, кси). В состав молекулы гемоглобина входят по две цепи двух разных типов.
Гем соединяется с белковой субъединицей, во-первых, через остаток гистидина координационной связью железа, во-вторых, через гидрофобные связи пиррольных колец и гидрофобных аминокислот. Гем располагается как бы "в кармане" своей цепи и формируется гемсодержащий протомер.
Нормальные формы гемоглобина
Существует несколько нормальных вариантов гемоглобина:
HbР – примитивный гемоглобин, содержит 2ξ- и 2ε-цепи, встречается в эмбрионе между 7-12 неделями жизни,
HbF – фетальный гемоглобин, содержит 2α- и 2γ-цепи, появляется через 12 недель внутриутробного развития и является основным после 3 месяцев,
HbA – гемоглобин взрослых, доля составляет 98%, содержит 2α- и 2β-цепи, у плода появляется через 3 месяца жизни и к рождению составляет 80% всего гемоглобина,
HbA2 – гемоглобин взрослых, доля составляет 2%, содержит 2α- и 2δ-цепи,
HbO2 – оксигемоглобин, образуется при связывании кислорода в легких, в легочных венах его 94-98% от всего количества гемоглобина,
HbCO2 – карбогемоглобин, образуется при связывании углекислого газа в тканях, в венозной крови составляет 15-20% от всего количества гемоглобина.
Патологические формы гемоглобина
HbS – гемоглобин серповидно-клеточной анемии.
MetHb – метгемоглобин, форма гемоглобина, включающая трехвалентный ион железа вместо двухвалентного. Такая форма обычно образуется спонтанно, в этом случае ферментативных мощностей клетки хватает на его восстановление. При использовании сульфаниламидов, употреблении нитрита натрия и нитратов пищевых продуктов, при недостаточности аскорбиновой кислоты ускоряется переход Fe2+ в Fe3+. Образующийся metHb не способен связывать кислород и возникает гипоксия тканей. Для восстановления ионов железа в клинике используют аскорбиновую кислоту и метиленовую синь.
Hb-CO – карбоксигемоглобин, образуется при наличии СО (угарный газ) во вдыхаемом воздухе. Он постоянно присутствует в крови в малых концентрациях, но его доля может колебаться от условий и образа жизни.
Угарный газ является активным ингибитором гем-содержащих ферментов, в частности, цитохромоксидазы 4-го комплекса дыхательной цепи.
HbA1С – гликозилированный гемоглобин. Концентрация его нарастает при хронической гипергликемии и является хорошим скрининговым показателем уровня глюкозы крови за длительный период времени.
№24.Производные гемоглобина.
В соответствии с функциональным назначением гемоглобина в физиологических условиях в организме образуются такие его производные, как: оксигемоглобин, карбгемогло-бин, и в следовых количествах — карбоксигемоглобин и метгемоглобин.
Оксигемоглобин (НbO2) образуется при обогащении крови кислородом в легких, за счет присоединения кислорода к гему гемоглобина при помощи координационных связей железа без изменения валентности железа.
Оксигемоглобин очень лабильное соединение и, поступая с кровью к тканям, где давление кислорода значительно ниже, чем в легких, он легко диссоциирует на свободный гемоглобин и кислород. Кислород утилизируется в процессе клеточного дыхания, а гемоглобин вступает в соединение с углекислым газом, образуя карбгемоглобин (НbСO2). Это также непрочное соединение, которое легко отдает углекислый газ при поступлении крови в легкие. Связь с углекислым газом образует не гем, а глобин за счет свободных аминогрупп аминокислот.
Карбоксигемоглобин (НbСО) — это соединение гемоглобина с окисью углерода. Образуется в результате диссимиляции гемоглобина и в норме не превышает 2% от общей массы гемоглобина. Карбоксигемоглобин, в отличие от других производных гемоглобина, является очень прочным соединением. СО образует ковалентную связь с гемоглобином, блокируя ассоциацию с ним кислорода. Сродство СО к гемоглобину более чем в 200 раз выше сродства кислорода. При отравлении угарным газом (СО), когда образуются значительные количества карбоксигемоглобина, гемоглобин теряет способность связывать кислород. Кроме того, СО способен попадать в клетки и ингибировать дыхательную цепь в митохондриях. Все это приводит к тканевой гипоксии, ацидозу, депрессии центральной нервной системы, блокаде дыхательного центра и наступает смерть от удушья.
Метгемоглобин (МетНb)—образуется при окислении двухвалентного железа гема в трехвалентное. При этом отмечается и переход окраски из красной в коричнево-зеленоватую. В норме метгемоглобин образуется за счет процессов аутоокисления и не превышает 2% от общей массы гемоглобина, так как небольшое количество образуемого трехвалентного железа до двухвалентного восстанавливает фермент метгемоглобин-редуктаза. Этот фермент позволяет поддерживать уровень MetHb в крови на очень низких цифрах.
При патологии количество МетНb в крови возрастает. Развивается состояние метгемоглобинемии, которое может быть следствием генетической патологии — генетически обусловленным дефицитом MetHb-редуктазы, либо носить приобретенный характер. Например, при отравлении окислами азота, цианидами и др. Образующиеся значительные количества метгемоглобина могут вызвать смерть от недостатка кислорода, так как метгемоглобин лишен способности связываться с кислородом.
№25,26.Липопротеины.
Липопротеи́ны (липопротеиды) — класс сложных белков, простетическая группа которых представлена каким-либо липидом. Так, в составе липопротеинов могут быть свободные жирные кислоты, нейтральные жиры, фосфолипиды, холестериды.
Липопротеины представляют собой комплексы, состоящие из белков (аполипопротеинов; сокращенно — апо-ЛП) и липидов, связь между которыми осуществляется посредством гидрофобных и электростатических взаимодействий.
Липопротеины подразделяют на свободные, или растворимые в воде (липопротеины плазмы крови, молока и др.), и нерастворимые, т. н. структурные (липопротеины мембран клетки, миелиновой оболочки нервных волокон, хлоропластов растений).
Среди свободных липопротеинов (они занимают ключевое положение в транспорте и метаболизме липидов) наиболее изучены липопротеины плазмы крови, которые классифицируют по их плотности. Чем выше содержание в них липидов, тем ниже плотность липопротеинов. Различают липопротеины очень низкой плотности (ЛПОНП), низкой плотности (ЛПНП), высокой плотности (ЛПВП) и хиломикроны. Каждая группа липопротеинов очень неоднородна по размерам частиц (наиболее крупные — хиломикроны) и содержанию в ней апо-липопротеинов. Все группы липопротеинов плазмы содержат полярные и неполярные липиды в разных соотношениях.
Виды липопротеиновВид Размеры Функция
Липопротеины высокой плотности (ЛВП) 8-11 нм Транспорт холестерина от периферийных тканей к печени
Липопротеины низкой плотности (ЛНП) 18-26 нм Транспорт холестерина, триацилглицеридов и фосфолипидов от печени к периферийным тканям
Липопротеины промежуточной (средней) плотности ЛПП (ЛСП) 25-35 нм Транспорт холестерина, триацилглицеридов и фосфолипидов от печени к периферийным тканям
Липопротеины очень низкой плотности (ЛОНП) 30-80 нм Транспорт холестерина, триацилглицеридов и фосфолипидов от печени к периферийным тканям
Хиломикроны 75-1200 нм Транспорт холестерина и жирных кислот, поступающих с пищей, из кишечника в периферические ткани и печень
Нековалентная связь в липопротеинах между белками и липидами имеет важное биологическое значение. Она обусловливает возможность свободного обмена липидов и модуляцию свойств липопротеинов в организме.
Липопротеины являются:
структурными элементами мембран клеток животных организмов;
транспортными белками, транспортирующими холестерин и другие стероиды, фосфолипиды и др.
№27.Биологические мембраны.
Кле́точная мембра́на (или цитолемма, или плазмалемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды.
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7—8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
Функции
барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
