
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Ø фаза быстрой реполяризации, обусловлена быстрым выходящим потенциалзависимым калиевым током, в период этой фазы медленные кальциевые каналы постепенно инактивируются, что ускоряет процесс реполяризации мембраны (поскольку резко ослабевает входящий кальциевый ток, деполяризующий мембрану). Реполяризация мембраны сопровождается постепенным закрытием потенциалзависимых калиевых каналов (полное их закрытие может длиться до 1с и поэтому, как правило, продолжается еще и в начале каждого следующего ПД) и реактивацией быстрых натриевых каналов (их возвратом к исходному состоянию, характерному для покоя), что приводит к постепенному восстановлению возбудимости миокардиальной клетки. В связи с отмеченным в эту фазу миокардиоцит находится в состоянии относительной рефрактерности.
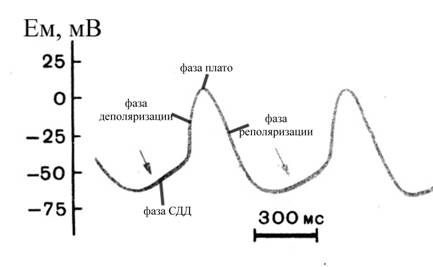 Реполяризация мембраны рабочего кардиомиоцита приводит к восстановлению МП до уровня, характерного для состояния покоя (-90мВ), и клетка продолжает пребывать в этом состоянии до момента прихода очередного нервного импульса. Амплитуда потенциала действия рабочего кардиомиоцита составляет около 120 мВ (реверсия 30мВ).
Реполяризация мембраны рабочего кардиомиоцита приводит к восстановлению МП до уровня, характерного для состояния покоя (-90мВ), и клетка продолжает пребывать в этом состоянии до момента прихода очередного нервного импульса. Амплитуда потенциала действия рабочего кардиомиоцита составляет около 120 мВ (реверсия 30мВ).
Рис. 14. Схема потенциала действия проводящего кардиомиоцита
Потенциал действия проводящих кардиомиоцитов характеризуется рядом отличительных черт от такового в рабочих клетках. Во-первых, он имеет гораздо меньшую амплитуду (50-60мВ, реверсия составляет 0÷+10мВ). Во-вторых, фаза его начальной деполяризации характеризуется меньшей крутизной нарастания, а фаза плато ПД – гораздо меньшей длительностью по сравнению с таковыми в рабочей миокардиальной клетке. В потенциале действия проводящих кардиомиоцитов выделяют следующие фазы:
Ø фаза спонтанной диастолической деполяризации, возникает вскоре после возврата МП к исходному уровню после предшествующего возбуждения, обусловлена
ü входящим натриевым током утечки (мембрана проводящих кардиомиоцитов в покое хорошо проницаема для натрия)
ü отчасти быстрым входящим потенциалзависимым натриевым током (потенциалзависимые натриевые каналы в пейсмекерных клетках в большинстве своем инактивируются при достижении мембранным потенциалом уровня -40мВ, в связи с чем принимают участие в СДД лишь в ее начале)
ü некоторым уменьшением калиевой проницаемости мембраны. В частности, потенциалзависимые калиевые каналы, обеспечивающие реполяризацию мембраны кардиомиоцита, характеризуются очень длительной инаткивацией, которая продолжается уже после ПД в момент СДД следующего ПД. Постепенное же полное закрытие этих каналов приводит к резкому ослаблению выходящего потенциалзависимого калиевого тока, что также способствует некоторой деполяризации мембраны кардиомиоцита в момент начинающейся очередной СДД
ü отчасти медленным входящим потенциалзависимым кальциевым током (включается во время последней 1/3 - 1/4 СДД). Потенциалзависимые медленные кальциевые каналы начинают активироваться деполяризацией при МП -40мВ, они гораздо хуже, но все же проницаемы и для ионов натрия
Ø фаза деполяризации, обусловлена медленным входящим потенциалзависимым кальциевым током (гораздо более медленным, чем потенциалзависимый натриевый ток), в связи с чем отличается гораздо меньшей крутизной нарастания и большей длительностью, чем соответствующая фаза в рабочих кардиомиоцитах, а также меньшей амплитудой. Причем для полного восстановления от инактивации потенциалзависимых медленных кальциевых каналов требуется несколько сотен мс, и реактивация кальциевых каналов продолжается даже после завершения ПД (уже в начале фазы СДД следующего ПД)
Ø фаза плато ПД, обусловлена продолжающимся, но гораздо более слабым, чем в предыдущей фазе, медленным потенциалзависимым кальциевым током. Уже во время предшествующей фазы деполяризации проводящих кардиомиоцитов медленные кальциевые каналы начинают инактивироваться (при МП около -20мВ), но поскольку их инактивация происходит очень медленно, то еще в течение значительного времени часть этих каналов остается открытой, обеспечивая фазу плато ПД. Данная фаза в проводящих кардиомиоцитах отличается гораздо меньшей продолжительностью по сравнению с соответствующей фазой в рабочем миокардиоците
Ø фаза быстрой реполяризации, подобно таковой в рабочих кардиомиоцитах обусловлена выходящим потенциалзависимым калиевым током; по ходу этой фазы возбудимость проводящих кардиомиоцитов постепенно восстанавливается, а по достижении мембранным потенциалом величины -50 ÷ -60 мВ, начинается новая СДД.
Влияние вегетативных нейромедиаторов (катехоламинов и ацетилхолина) на электрическую активность пейсмекерных клеток
Катехоламины через посредство b1-адренорецепторов пейсмекерных клеток синоатриального узла оказывают на них следующие эффекты:
Ø увеличивают амплитуду потенциалзависимого медленного входящего кальциевого тока без изменений его кинетики, что способствует, во-первых, ускорению СДД, а во-вторых, повышению крутизны фазы деполяризации ПД, которое, в свою очередь, необходимо для увеличения надежности и скорости проведения возбуждения в синоатриальном узле;
Ø увеличивают амплитуду потенциалзависимого выходящего калиевого тока без изменений его кинетики, что способствует ускорению фазы реполяризации, а следовательно, укорочению ПД в пейсмекерных клетках и учащению их разрядов
Ø изменяют кинетику потенциалзависимого быстрого входящего натриевого тока, «заставляя» потенциалзависимые натриевые каналы работать при менее отрицательных значениях МП (т. е. при таких значениях, при которых обычно эти каналы инактивированы). Сдвигая порог активации быстрых потенциалзависимых натриевых каналов в сторону более положительных мембранных потенциалов, катехоламины делают тем самым их работающими, что продлевает существование быстрого входящего натриевого тока в момент СДД, таким образом ускоряя ее.
Ацетилхолин через посредство М2-холинорецепторов пейсмекерных клеток оказывает на них следующее действие:
Ø угнетает потенциалзависимый медленный входящий кальциевый ток, что приводит к ослаблению и удлинению СДД, а также уменьшению амплитуды и длительности ПД (за счет фазы деполяризации ПД)
Ø способствует открытию в мембране пейсмекеров специальных каналов, чувствительных к ацетилхолину (хемовозбудимых каналов), которые проницаемы для калия. Усиленный же выход калия из клетки будет прямо противоположным образом сказываться на длительности сердечного цикла в зависимости от того, на какую фазу ПД он приходится (фазозависимость эффекта ацетилхолина на пейсмекерные клетки). Так, усиление выхода калия из пейсмекерной клетки в момент фазы СДД способствует гиперполяризации мембраны и замедляет СДД (что повлечет за собой урежение сердечной деятельности), тогда как открытие этих каналов в момент фазы плато ПД и реполяризации, напротив, будет укорачивать ПД, способствуя учащению сердечного ритма.
При длительном действии ацетилхолина на пейсмекерные клетки реализуются механизмы, способствующие ускользанию сердца из-под влияния ацетилхолина и соответственно блуждающего нерва, в основе которых лежат
Ø постепенная десенситизация холинорецепторов,
Ø активация вследствие гиперполяризации мембраны быстрого потенциалзависимого натриевого тока, который будет воспрепятствовать такой гиперпоялризации.
6. Понятие о сердечном цикле, его фазы
Сердечный цикл – это период деятельности сердца, включающий одно сокращение его камер и последующее их расслабление (общую паузу). Сердечный цикл складывается из:
Ø сокращения предсердий
Ø сокращения желудочков
Ø общей паузы, при которой и желудочки и предсердия расслаблены.
Продолжительность сердечного цикла у человека при частоте сердечных сокращений 75 уд/мин составляет 0,8с. В сердечном цикле выделяют периоды (основные этапы систолы или диастолы), фазы (части того или иного периода) и интервалы (переходные межфазовые состояния).
Сердечный цикл имеет следующую структуру:
Ø систола предсердий (является началом каждого цикла, длится около 0,1с, охват возбуждением и последующее сокращение миокардиоцитов предсердий происходит асинхронно и поэтому давление в них повышается от нуля всего до 5-8 мм. рт. ст.; кровь пассивно из предсердий поступает в расслабленные желудочки, при этом вернуться обратно в вены она не может, поскольку первыми в предсердиях возбуждаются и сокращаются мышечный волокна, окружающие устья вен)
Ø систола желудочков, длиться около 0,33с, разделяется на несколько периодов:
ü период напряжения, в ходе которого рабочие кардиомиоциты желудочков охватываются возбуждением и последующим сокращением, давление в полостях желудочков постепенно нарастает и в момент, когда оно становиться выше такового в аорте и легочной артерии, начинается следующий период (изгнание крови желудочками). Средняя длительность периода напряжения составляет 0,08с; он включает две фазы:
§ фаза асинхронного сокращения кардиомиоцитов желудочков, длится около 0,05с, начало этой фазы совпадает с началом деполяризации рабочего миокарда желудочков и соответствует моменту появления зубца Q на ЭКГ. Во время этой фазы возбуждение и последующее сокращение постепенно распространяются по рабочему миокарду желудочков, в результате чего давление в их полостях очень медленно растет, а к концу этой фазы, когда почти все кардиомиоциты переходят к сокращению, начинает быстро нарастать, и кровь устремляется из желудочков в направлении предсердий (в которых давление почти равно нулю), захлопывая атриовентрикулярные клапаны. В момент захлопывания атриовентрикулярных клапанов возникает I или систолический тон сердца. Именно с момента захлопывания атриовентрикулярных клапанов начинается следующая фаза периода напряжения желудочков
§ фаза изометрического сокращения кардиомиоцитов желудочков, длиться около 0,03с. В момент этой фазы уже все рабочие кардиомиоциты вовлечены в сократительный акт, атриовентрикулярные клапаны уже закрыты, полулунные – еще остаются закрытыми, объем крови в полостях желудочков является постоянным, в результате чего сердечные мышечные волокна сокращаются в изометрическом режиме (т. е. при сокращении длина их остается постоянной, а они развивают лишь напряжение). Давление крови в желудочках резко нарастает: до 70-80мм. рт. ст. в левом и до 15-20мм. рт. ст. в правом. Левый желудочек приобретает круглую форму и с силой ударяет о внутреннюю поверхность грудной стенки, в результате чего в пятом межреберье, на 1см слева от среднеключичной линии ощущается сердечный толчок. К концу этого периода давление в полостях желудочков становится выше такового в соответствующих артериях, выносящих кровь из сердца, что делает возможным последующий выброс желудочками порции крови в артериальные сосуды. Как следствие, на смену этой фазе приходит следующий период – период изгнания крови желудочками
ü период изгнания крови из желудочков, длится около 0,25с, начинается с открытия кровью, движущейся в направлении артерий, полулунных клапанов. В этот период рабочие кардиомиоциты желудочков продолжают сокращаться, вследствие чего давление в их полостях продолжает нарастать (до 120-130 мм. рт. ст. в левом и до 25мм. рт. ст. в правом). Период изгнания состоит из:
§ протосфигмического интервала (0,005с), представляющего собой межфазовое состояние, соответствующее моменту открытия полулунных клапанов. Выброс же крови из сердца начинается не тотчас после достижения в желудочках давления, превышающего таковое в аорте и легочной артерии, а лишь после того, как будет преодолена инерция столба крови, заполняющей эти сосуды
§ фазы быстрого или максимального изгнания (0,12с), во время которой сердечные мышечные волокна в большинстве своем еще сокращаются, и давление крови в полостях желудочков продолжает нарастать, в результате чего она быстро изливается в аорту и легочную артерию, а объем желудочков резко уменьшается; именно в эту фазу выбрасывается большая часть систолического объема крови
§ фазы медленного изгнания (0,13с), во время которой сокращения миокарда желудочков постепенно ослабевает вследствие постепенного начала расслабления его рабочих кардиомиоцитов, и кровь в артерии изливается гораздо медленнее по сравнению с предыдущей фазой. Кроме того, в эту фазу отток крови из аорты и легочного ствола к периферии начинает превышать ее поступление из сердца, и систолическое давление в этих сосудах начинает постепенно снижаться
В момент, когда большая часть миокардиоцитов желудочков начинает расслабляться, давление в них резко падает, и выброс крови прекращается, начинается диастола желудочков (длится около 0.47с), включающая следующие периоды:
ü протодиастолический период (0,04с), представляет собой переходное межфазовое состояние, соответствующее времени, затрачиваемому на закрытие полулунных клапанов кровью, движущейся из аорты и легочного ствола в направлении расслабляющихся желудочков. В момент захлопывания полулунных клапанов возникает II или диастолический тон сердца
ü период изометрического (изоволюметрического) расслабления желудочков (0,08с). Начинается от момента захлопывания полулунных клапанов; в этот период атриовентрикулярные клапаны еще остаются закрытыми, и миокард желудочков расслабляется с постоянным объемом крови, оставшейся в их полостях после систолы. В связи с тем, что расслабление желудочков происходит при постоянном объеме крови в них, эту фазу называют изометрической. К концу этого периода давление в полостях желудочков становится чуть ниже такового в предсердиях, что приводит к открытию движущейся из предсердий кровью атриовентрикулярных клапанов и началу следующего периода
ü период наполнения желудочков кровью (0,25с). В этот период и предсердия и желудочки расслаблены, и кровь движется пассивно из предсердий (которые принимают ее из вен) в желудочки; причем во время этого периода в желудочки притекает около 70% от общего венозного возврата крови к сердцу. Данный период включает две фазы:
§ фаза быстрого наполнения (0,08с), начинается одновременно с открытием створчатых клапанов. Во время этой фазы происходит основное по объему наполнение желудочков кровью. Колебания стенок желудочков вследствие быстрого притока крови к ним вызывают появление III тона сердца
§ фаза медленного наполнения или диастазис (0,17с), во время которой существенного наполнения желудочков кровью не происходит; почти полностью редуцируется при частоте сердечных сокращений свыше 110-130уд/мин; при еще более частом ритме резко сокращается и фаза быстрого наполнения
ü пресистоличсекий период, соответствующий систоле предсердий (0,1с). По окончании фазы медленного наполнения желудочков кровью начинается очередная систола предсердий, знаменующая начало следующего сердечного цикла, и происходит дополнительная подкачка небольшого количества крови в желудочки (около 30% от общего венозного возврата крови к сердцу). Колебание стенок сердца, вызванное сокращением предсердий и дополнительным притоком крови в желудочки, обуславливает возникновение IV тона сердца. Причем при обычной аускультации слышны только первые два тона (систолический и диастолический), тогда как III и IV тоны являются тихими и выявляются только при графической их регистрации
ü интерсистолический интервал (0,007с) представляет собой время между окончанием систолы предсердий и началом систолы желудочков; он не является постоянной составной частью сердечного цикла.
Периоды сердечного цикла от начала расслабления желудочков до момента наступления систолы предсердий очередного сердечного цикла составляют в своей совокупности общую паузу сердца, включающую таким образом протодиастолический интервал, период изометрического расслабления желудочков и период наполнения желудочков кровью.
Большинство фаз деятельности правого и левого желудочков начинается не строго одновременно, что связано с различными экстракардиальными условиями для правого и левого отделов сердца (в частности, правый желудочек осуществляет выброс крови против меньшего сопротивления, чем левый) и неодинаковыми сократительными свойствами миокарда обоих желудочков. Так, фаза асинхронного сокращения правого желудочка длительнее, чем левого, а фаза изометрического сокращения и период изгнания крови, составляющие в совокупности механическую систолу желудочков, в левом желудочке начинаются раньше, чем в правом; изгнание же крови из правого желудочка начинается, как правило, на сотые доли секунды раньше, чем из левого.
Учащение и урежение сердечного ритма определенным образом отражаются на продолжительности фаз сердечного цикла. Так, учащение сердечной деятельности приводит к укорочению периода изгнания, механической и общей систолы, диастазиса и систолы предсердий, тогда как урежение – сопровождается противоположными фазовыми сдвигами.
7. Механизмы венозного возврата крови к сердцу
Кровь как в самом сердце (из одних его камер в другие), так и в сосудистом русле движется пассивно по градиенту давления (из области с более высоким давлением в область с более низким давлением). Обратный возврат крови из сосудистого руса в сердце (т. н. венозный возврат) возможен благодаря следующим механизмам:
Ø остаточной движущей силе сердца – той части общей энергии, создаваемой деятельностью сердца как насоса, которой обладает движущаяся кровь в венозной отделе сосудистого русла. Дело в том, что сокращающееся сердце сообщает изгоняемой в сосудистое русло порции крови определенный запас энергии, которая большей частью расходуется на преодоление сил сопротивления при движении крови по сосудам. Между тем кровь при своем движении расходует эту энергии не полностью, а небольшая ее доля (соответствующая к примеру в большом круге кровообращения давлению в 7мм. рт. ст) все же сохраняется у венозной крови, заполняющей полые и легочные вены. Давление же в полостях сердца в момент их диастолы почти равно нулю. Таким образом, создается очень небольшой градиент давления между венами, возвращающими кровь в сердце, и полостями сердца в момент диастолы, который отчасти облегчает венозный возврат крови к сердцу
Ø работе венозной помпы, представляющей собой совокупность работающих в каждый данный момент скелетных мышц, сокращение которых приводит к наружному сдавливанию вен конечностей и туловища, в большинстве из которых имеются клапаны, пропускающие кровь только в одном направлении (к сердцу)
Ø присасывающему действию грудной клетки или работе торако-абдоминального насоса, сущность которого состоит в следующем. В момент вдоха грудная клетка увеличивается в объеме и давление в ее органах (в том числе в грудной части полых вен) в результате их некоторого растяжения уменьшается, тогда как объем брюшной полости во время вдоха, наоборот, уменьшается, а давление в ее органах (в том числе брюшной части полых вен) несколько возрастает. Таким образом, при вдохе увеличивается градиент давления между грудной и брюшной частями нижней полой вены, а также несколько понижается давление в грудной части верхней полой вены, что облегчает приток крови к ним
Ø присасывающего действия самого сердца, сущность которого состоит в следующем. В момент систолы желудочков, сопровождающейся укорочением их вертикального размера, предсердно-желудочковая перегородка оттягивается книзу, что приводит к расширению полостей предсердий и облегчению притока крови к ним из вен.
8. Систолический и минутный объем кровотока как основные параметры интенсивности сердечной деятельности
Ключевыми параметрами, отражающими, с одной стороны, интенсивность сердечной деятельности, а с другой – функциональный резерв сердечно сосудистой системы, являются систолический и минутный объем кровотока.
Систолический (ударный) объем (выброс) – это количество крови, выбрасываемое одним из желудочков (левым или правым) за одну систолу. В норме его величина является одинаковой для правого и левого желудочков и составляет 60-75мл. Большая же толщина стенки левого желудочка, по сравнению с таковой правого, связана с тем, что левый желудочек за каждую систолу изгоняет тот же объем крови, что и правый, но против большего сопротивления выбросу (давление в аорте в момент диастолы составляет 70-80мм. рт. ст., а в легочном стволе – всего 10-15мм. рт. ст.). Как правило, за каждую систолу желудочки изгоняют от 1/3 до 1/2 всего имеющегося в них объема крови (т. е. того объема, который притекает к желудочкам в момент диастолы – конечно-диастолического объема). То же количество крови, которое остается в желудочках после их систолы составляет их остаточный (резервный или резедуальный) объем. Именно за счет этого остаточного объема и может быть увеличен систолический выброс в случае необходимости (например, при тяжелой мышечной работе, эмоциональных состояниях, когда усиливаются инотропные влияния на миокард). Следовательно, величина остаточного объема крови, во многом зависящая от диастолического объема желудочков, отчасти косвенно характеризует функциональный резерв сердца, определяемый степенью максимально возможного увеличения его нагнетательной функции в случае необходимости. А именно, чем больше конечно-диастолический объем желудочков, тем соответственно потенциально и больше крови может выбрасываться ими в артериальную часть сосудистого русла в случае необходимости (т. е. тем выше функциональный резерв сердца). Вторым фактором, определяющим функциональный резерв сердца, является максимально возможная сила сердечных сокращений.
Таким образом, величина систолического выброса определяется следующими параметрами:
Ø конечно-диастолическим объемом желудочков сердца, зависящим, в свою очередь, от венозного возврата крови к сердцу в момент диастолы и максимально возможного диастолического размера камер сердца
Ø силы сокращения желудочков.
У разных людей функциональный резерв сердца может значительно отличаться. Так, у большинства спортсменов, как правило, наблюдается физиологическая дилятация (расширение) полостей сердца и некоторая гипертрофия миокарда желудочков, в результате чего оказываются увеличенными, по сравнению с нетренированными людьми, как максимально возможный конечно-диастолический размер желудочков (а значит, и конечно-диастолический объем крови), так и сила сердечных сокращений. У людей, страдающих сердечной недостаточностью, также наблюдается некоторая дилятация полостей сердца, но сила сердечных сокращений снижена, по сравнению с нормальными людьми, в результате чего систолический выброс, как в покое, так и при выполнении каких-либо физических нагрузок, оказывается меньше значений здоровых людей.
Минутный объем сердца (или минутный объем кровотока) – это количество крови, выбрасываемое каким-то из желудочков сердца за одну минуту; соответственно это же количество крови проходит через суммарное поперечное сечение любого участка большого или малого круга кровообращения за одну минуту. Минутный объем кровотока определяется как произведение систолического выброса на частоту сердечных сокращений (т. е. МО = СО·ЧСС). В норме в покое его величина у взрослого человека составляет от 4 до 5 л/мин. Таким образом, минутный объем кровотока зависит от:
Ø величины систолического выброса, находящейся в зависимости от:
ü венозного возврата крови к сердцу,
ü конечно-диастолического размера желудочков
ü силы их сокращений
Ø частоты сердечных сокращений.
Именно величина минутного объема кровотока является фактором, лимитирующим уровень кровоснабжения всех органов организма, в том числе и самого сердца. Следовательно, минутный объем кровотока является интегральным показателем, зависящим, с одной стороны, от интенсивности работы сердца (частоты и силы сердечных сокращений), а с другой – от потребности периферических тканей в уровне кровоснабжения. Максимально возможная степень увеличения минутного объема в случае необходимости характеризует функциональный резерв аппарата кровообращения. Увеличение минутного объема кровотока может достигаться, как минимум, двумя путями: за счет роста частоты сердечных сокращений и величины систолического выброса. У здоровых людей в случае необходимости (при физической нагрузке, эмоциональных состояниях) частота сердечных сокращений может возрасти в 2-3 раза, систолический выброс – в 1,5-2 раза, а минутный объем кровотока – 5-6 раза. Причем слишком значительное увеличение частоты сердечных сокращений (свыше 180 уд/мин) является неэффективным в плане увеличения минутного объема кровотока, потому что приводит к резкому сокращению диастолы, а значит, и нарушению нормального наполнения сердца кровью, что негативно отражается на последующей его систоле. Немаловажное значение для нормального кровоснабжения органов, интенсивно работающих при различных физиологических состояниях, имеет не только увеличение минутного объема кровотока, но и перераспределение крови между органами, работающими с разной интенсивностью. В частности, артериолы усиленно работающих в каждый данный момент органов максимально расширяются, а органов, пребывающих в состоянии относительного покоя – напротив, суживаются (перераспределительный механизм), в результате чего на фоне некоторого общего повышения минутного объема кровотока резко увеличивается доставка крови к наиболее интенсивно функционирующим органам (т. е. возрастает их регионарный кровоток).
9. Механизмы регуляции сердечной деятельности
Сердце относится к органам, работающим на протяжении жизни постоянно в ритмическом режиме. Причем оно очень чутко реагирует на изменяющиеся при определенных физиологических состояниях потребности различных периферических тканей в уровне кровоснабжения. В норме (при отсутствии патологических изменений в сердечно-сосудистой и дыхательной системах) интенсивность работы сердца в каждый момент жизнедеятельности является строго адекватной потребностям всех органов организма (в том числе самого сердца) в уровне кровоснабжения, который определяет доставку к тканям кислорода и питательных веществ. Такое строгое соответствие между эффективностью работы сердца и потребностью организма в кровоснабжении, являющееся залогом нормальной жизнедеятельности, достигается благодаря разнообразным механизмам регуляции сердечной деятельности. Все существующие механизмы регуляции работы сердца в первом приближении можно классифицировать следующим образом:
Ø внутрисердечные, включающие
ü внутриклеточные механизмы регуляции. Сущность их состоит в том, что уровень обменных процессов в кардиомиоцитах, синтез ключевых ферментов и структурных белков (в том числе белков миофибрилл) в них напрямую зависит от интенсивности функционирования. В частности, физиологические состояния, сопровождающиеся продолжительной интенсификацией сердечной деятельности, приводят, как правило, к рабочей гипертрофии кардиомиоцитов и в целом определенных камер сердца в результате усиления синтеза структурных белков в них. Как следствие, гипертрофированные кардиомиоциты могут выполнять больший объем работы, чем нормальные, и повышенная функциональная активность сердечной мышцы получает свое морфологическое обеспечение (т. н. рабочая гипертрофия миокарда в ответ на развиваемую им гиперфункцию). Кроме того, внутриклеточные механизмы регуляции сердечной деятельности могут проявляться в изменении силы сокращения кардиомиоцитов в зависимости от степени их растяжения в момент диастолы (гетерометрический механизм регуляции сердечной деятельности или закон Франка-Старлинга). Сущность этого закона состоит в том, что, чем больше крови притекает к сердцу в момент диастолы (т. е. чем сильнее растягивается сердечная мышца и соответственно ее кардиомиоциты), тем больше резервных мостиков образуется в миофибриллярном аппарате (подобно таковому в умеренно растянутой скелетной мышце) и тем больше будет сила сокращения миокарда в момент последующей систолы. Однако такой механизм реализуется только при пониженном или в крайнем случае близком к нормальному давлению крови в артериальном русле, тогда как при повышенном артериальном давлении его реализации препятствуют внесердечные рефлексы, направленные на нормализацию артериального давления и замедляющие и ослабляющие работу сердца. Наряду с описанной положительной зависимостью силы сердечных сокращений от исходной длины кардиомиоцитов в момент диастолы, существует и позитивная связь между силой сокращения кардиомиоцитов и давлением в начальной части артериального русла (т. н. сопротивлением выбросу). Механизм регуляции сердечной деятельности, лежащий в основе этой зависимости носит название гомеометрического механизма регуляции сердца (раскрыт Анрепом) и во многом вытекает из закона Фрнака-Старлинга. Так, повышение давления в начальном отделе сосудистого русла (повышение сопротивления выбросу) приводит к тому, что первоначально при каждой систоле объем выбрасываемой крови несколько снижается (поскольку желудочкам приходится работать против большего сопротивления выбросу), а следовательно, остаточный объем крови возрастает. Отмеченное приводит к тому, что в условиях повышенного сопротивления выбросу после каждой систолы в полостях сердца накапливается все большее и большее количество крови. Это обуславливает некоторое перерастяжение кардиомиоцитов и соответственно большую силу их последующих сокращений. Гомеометрический механизм регуляции сердечной деятельности, подобно гетерометрическому, реализуется только в условиях пониженного системного артериального давления.
ü регуляцию деятельности сердца посредством межклеточных взаимодействий в нем. Такая регуляция достигается благодаря тесным структурным и функциональным связям как в пределах популяции кардиомиоцитов, так и кардиомиоцитов с соединительнотканными клетками, обеспечивающими трофику сердечных мышечных. Так, нормальное проведение возбуждения в миокарде и соответственно ритмичная его работа возможны только в случае нормального состояния нексусов, обеспечивающих передачу нервного импульса с одного кардиомиоцита на соседние. Соединительнотканные клетки, наряду с тем, что транспортируют питательные вещества в кардиомиоциты из крови, способны вырабатывать и определенные факторы (белковой природы), влияющие на экспрессию генетической информации в сердечных мышечных клетках, в том числе, на синтез структурных белков в них.
ü внутрисердечные периферические рефлексы, замыкаются на уровне внутрисердечных (интрамуральных) парасимпатических ганглиев, формирующих в совокупности метасимпатическую систему. Эфферентные нейроны интрамуральных парасимпатических ганглиев сердца выступают в роли конечного эфферентного пути как для информации, поступающей из ц. н.с., так и проводимой от рецепторов самого сердца, коронарных сосудов, начальных отделов аорты и легочного ствола и дистальных отделов вен, впадающих в сердце. На уровне этих ганглиев замыкаются короткие (периферические) дуги рефлексов, способных изменять сердечную деятельность в зависимости от его текущего состояния. Отличительной особенностью этих рефлексов является то, что они возникают в ответ на возбуждение рецепторов сердца, коронарных сосудов, начальных отделов артериального русла или венозных сосудов, впадающих в сердце, и состоят в изменении деятельности самого сердца. Примером таких рефлексов является рефлекс Бейнбриджа, проявляющийся в усилении сердечной деятельности (увеличении частоты и силы сердечных сокращений) в ответ на перерастяжение кровью полостей сердца и вен, впадающих в него. Данный рефлекс способствует перекачиванию большего объема крови в артериальное русло, тем самым препятствуя венозному застою. Однако реализуется он только в случае пониженного или нормального давления в сосудистом русле, тогда как его реализации при повышенном артериальном давлении препятствуют как другие внутрисердечные рефлексы (возникающие в ответ на возбуждение рецепторов растяжения коронарных сосудов и полостей сердца), так и внесердечные рефлексы (срабатывающие в ответ на повышение давления в магистральных артериях), направленные на нормализацию системного артериального давления. Примером внутрисердечных рефлексов может служить также увеличение силы сокращения не только левого, но и правого отделов сердца, возникающее в ответ на возбуждение рецепторов растяжения левого предсердия увеличенным объемом притекающей крови. Данный рефлекс способствует сочетанному изменению деятельности левой и правой половин сердца в случае раздражения рецепторов только какой-то одной из его половин, что подготавливает вторую половину к принятию большего объема крови и координированной работе левого и правого желудочков.
Ø внесердечные, к которым относятся:
ü нервные (обеспечиваются вегетативной нервной системой). Симпатические центры, регулирующие работу сердца (первые эфферентные симпатические нейроны), заложены в боковых рогах серого вещества верхних пяти грудных сегментов спинного мозга, а тела вторых эфферентных симпатических нейронов – в последних шейных и верхних грудных паравертебральных ганглиях. Аксоны же этих вторых эфферентных симпатических нейронов (постганглионарные симпатические волокна) непосредственно контактируют с проводящими и рабочими кардиомиоцитами, а также с гладкомышечными клетками сосудов сердца, не вступая во взаимодействие с внутрисердечной периферической нервной системой (метасимпатической нервной системой), образованной интрамуральными парасимпатическими узлами. В связи с отмеченным симпатический отдел оказывает на сердце всегда однонаправленные влияния, а именно увеличивает силу сердечных сокращений (положительный инотропный эффект), их частоту (положительный хронотропный эффект), повышает возбудимость (положительный батмотропный эффект) и проводимость (положительный дромотропный эффект) в сердечной мышце. Парасимпатический центр регуляции работы сердца заложен в продолговатом мозге и представлен скоплением первых эфферентных парасимпатических нейронов вегетативного ядра блуждающего нерва (заднее ядро блуждающего нерва), аксоны же этих нейронов в составе блуждающего нерва направляются к сердцу, где переключаются на вторые эфферентные парасимпатические нейроны, заложенные в интрамуральных ганглиях сердца. Наряду с эфферентными нейронами в интрамуральных ганглиях сердца имеются и чувствительные нейроны, дендриты которых образуют рецепторы в самом сердце и в коронарных сосудах, а аксоны могут переключаться на вторые эфферентные парсимптические нейроны (либо непосредственно, либо через вставочные нейроны интрамуральных узлов). Таким образом, вторые эфферентные парсимпатические нейроны сердечных интрамуральных ганглиев являются общим конечным путем для той информации, которая поступает из ц. н.с. (по волокнам блуждающего нерва), а также от рецепторов самого сердца и его сосудов. В связи с отмеченным парасимпатическая регуляция сердечной деятельности является более сложной и тонкой, чем симпатическая, и влияния, поступающие к сердцу по волокнам блуждающего нерва со стороны ц. н.с., могут отчасти модифицироваться в зависимости от текущего состояния сердечной мышцы. В целом, парасимпатический отдел нервной системы оказывает на сердце диаметрально противоположные таковым симпатического отдела эффекты (отрицательное ино-, хроно-, батмо - и дромотропное действие). Сильное возбуждение заднего ядра блуждающего нерва может в начале привести и к полной остановке сердца, но в дальнейшем, несмотря на продолжающееся тормозное действие блуждающего нерва на сердце, оно постепенно восстанавливает свою деятельность (ускользание сердца из под влияния блуждающего нерва). В основе такого ускользания лежат разнообразные механизмы, среди которых определяющую роль имеет десесинтизация холинорецепторов (снижение их сродства к ацетилхолину) и компенсаторные процессы в самих кардиомиоцитах, направленные на преодоление гиперполяризации мембраны. Медиатором в передаче симпатических влияний на миокард является норадреналин, а парасимпатических – ацетилхолин. Норадреналин, по сравнению с ацетилхолином, характеризуется большим латентным периодом действия на миокард (до 10с и более) и большей продолжительностью своего действия (в связи с более медленным разрушением определенными ферментами). Нейроны заднего ядра блуждающего нерва, имеющие отношение к регуляции сердечной деятельности, находятся в состоянии постоянного тонуса (некоторого базального возбуждения), который поддерживается постоянно поступающей к ним афферентной информацией от рецепторов сосудов (прессо - и хеморецепторов магистральных и периферических сосудов) и самого сердца (рецепторов растяжения и волюморецептров камер сердца), а также действием некоторых гуморальных факторов, циркулирующих в крови и церебральной жидкости. В связи с отмеченным парасимпатический отдел нервной системы оказывает постоянное тормозное влияние на миокард, а двусторонняя перерезка блуждающих нервов у млекопитающих животных и человека (кроме новорожденных особей) сопровождается учащением сердечной деятельности вследствие выключения такого тормозного влияния.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
