

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Предисловие.
Первоначально, статью задумывал с целью призвать читателя попробовать найти закономерность, которую называю целочисленностепенной, для связи «соседних» таксонов в технике, по аналогии с подобной закономерности в биологии, претендуя на утверждение о том, что она является одной из универсальных эволюционных закономерностей аналогично тому, как ранговая гиперболическая закономерность работает как в биологи (для биоценозов), так и в технике (в техноценозах) и более обще – в социально-экономических явлениях (например, распределение богатства).
Но смысл статьи выходит далеко за рамки этой первоначальной задумки.
Статья можно быть интересна не только (и не столько) с точки зрения теории биологической, технологической или социальной эволюций, но ещё больше – с точки зрения синтеза того и другого и третьего. В не малой степени благодаря тому, что в ней, на примерах подвожу к универсальной закономерности (верной для социальной и биологической эволюций) смены форм эволюции как следствие эффекта её ускоренного характера. Например, при переходе от биологической к технологической (социальной) эволюциям. Откуда можно пытаться делать выводы и относительно того, что нас ожидает.
В частности, в этом ключе (ключе ускоренного характера эволюции) рассматриваю причину появления человека как следствие того, что на определённом (и только на определённом) этапе эволюции, когда её ускоренность становится достаточно высокой, он должен возникнуть как неизбежная реакция на невозможность продолжения ускоренной эволюции с теми темпами, с которыми она шла лишь в рамках биологической формы. Поскольку вследствие ускоренного характера новые биологические формы просто не поспевают возникать за стремительным ростом новых жизненных ниш, которые и начинают заполняться человеком. Т. к. он, благодаря технологическим приспособлениям, способен заполнять их гораздо быстрее, чем возможные биологические аналоги, хоть и не столь эффективно.
24 октября 2013
Степенно-логарифмическая связь между количеством соседних таксонов в биологии как основа для поиска аналогичной связи в технике
Междисциплинарность ценологических представлений. Общая и прикладная ценология. Труды XIV конференции-семинара с международным участием по технетике и ценологии (Москва, 19 ноября 2009 г.) / Под общ. ред. проф. . Вып. 43. "Ценологические исследования". – М.: Технетика, 2010. – 256 с.
Введение.
Открытие гиперболических ранговых распределений технологических таксонов в техноценозах ознаменовало новый этап в развитии теории эволюции как технических (социальных) систем, так и биологических. Потому, что это поставило интригующий вопрос о единстве происхождения упомянутых распределений с ранее известными ранговыми гиперболическими распределениями таксонов в биологии. Основанная школа вскрыла не мало общих черт в развитии техноценозов и биоценозов. Наличие которых, видимо, не случайно и отражает единство каких-то фундаментальных принципов развития. В этом отношении эмпирические численные закономерности био - и техноценозов играют важную роль. Т. к. являются основой для понимания сути и построения единых, биолого-технологических математических моделей эволюции.
В этой статье, на основе биологического материала, я изложу и обосную ещё одну численную, эмпирическую закономерность, которая, предполагаю, свойственна не только для биологической эволюции, но и для технологической. Последнее ещё требует проверки. И основным мотивом написания этой статьи явилось стремление привлечь внимание к этому вопросу с целью, чтобы эта проверка была выполнена. Если гипотеза окажется верной, то в руках исследователей появится ещё один универсальный инструмент-критерий для построения структурных численных моделей теории эволюции (как социальной, так и биологической).
В статье так же изложен косвенный способ проверки упомянутой гипотезы применительно к технике.
Суть закономерности, о которой идёт речь, состоит в том, что степень от логарифма количества какой-либо категории таксонов ![]() , с точностью до константы, равна количеству надтаксонов
, с точностью до константы, равна количеству надтаксонов ![]() :
:
![]() (1)
(1)
Причём, как показывает анализ по биологическому материалу, если брать в качестве упомянутого логарифма логарифм натуральный, то коэффициент пропорциональности ![]() оказывается близким к единице или порядка того. А степень
оказывается близким к единице или порядка того. А степень ![]() оказывается близкой к показателю экспоненты. В случае биологии
оказывается близкой к показателю экспоненты. В случае биологии ![]() , например, может быть количеством родов, а
, например, может быть количеством родов, а ![]() - общим количеством семейств. Либо, если в качестве n взять количество семейств, то в качестве
- общим количеством семейств. Либо, если в качестве n взять количество семейств, то в качестве ![]() следует взять количество отрядов.
следует взять количество отрядов.
Что взять в качестве ![]() и
и ![]() в случае технических систем – вопрос не ясный и требует проработки. Но, думаю, что точно так же, как в биологии, естественным образом, выработалась линейка
в случае технических систем – вопрос не ясный и требует проработки. Но, думаю, что точно так же, как в биологии, естественным образом, выработалась линейка
«вид», «род», «семейство», «отряд», «класс», «тип»
в технических системах, аналогично, естественным образом тоже может быть выработана аналогичная линейка (для технических таксонов количество элементов в ней может быть другим). Такая, что соседние таксоны этой линейки удовлетворяют выше рассмотренной степенно-логарифмической связи. Не исключено, например, что ![]() может быть общим количеством видов всех изделий, а
может быть общим количеством видов всех изделий, а ![]() - общим количеством родов всех изделий которые существуют в данный момент.
- общим количеством родов всех изделий которые существуют в данный момент.
Задача проверки этой закономерности для видов и родов изделий, в настоящее время, скорее всего, непосильна в силу относительно быстрого роста количества этих таксонов и трудности их учёта. То же самое относится и к другим очень быстро растущим таксонам. Хотя эту задачу, вероятно, отчасти, можно упростить, рассматривая вместо общего количества таксонов ![]() и «соседних» надтаксонов
и «соседних» надтаксонов ![]() некоторую такую из них, относительно малую, (равновероятную) выборку
некоторую такую из них, относительно малую, (равновероятную) выборку ![]() ,
, ![]() , чтобы эта выборка была пропорциональна реальному количеству таксонов:
, чтобы эта выборка была пропорциональна реальному количеству таксонов:  ,
, 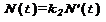 . Где
. Где ![]() ,
, ![]() – некоторые константы. Тогда, после подстановки последних двух соотношений в (1), получится соотношение с 4-мя константами, которые можно попытаться определить путём оптимизации по набору данных для эволюции
– некоторые константы. Тогда, после подстановки последних двух соотношений в (1), получится соотношение с 4-мя константами, которые можно попытаться определить путём оптимизации по набору данных для эволюции ![]() и
и ![]() .
.
Ещё одно возможное упрощение может состоять в выделении какие-то достаточно крупных таксонов (для них, вероятно, ещё не устоялись названия), количество которых относительно не велико и учёт которых технически возможен. Думаю, что имеет смысл начать с таксона - отрасль (например, авиастроение, машиностроение, ракетостроение, приборостроение, компьютерная отрасль, станкостроение, роботостроение и т. д.. и т. п.).
Скажем, первым этапом, за последние лет двести выявить даты возникновения (жизни) разных существующих (существовавших) отраслей.
Следующим этапом можно попытаться выявить существующее дробление внутри отраслей. Вероятно, что в каждой отрасли уже существуют какие-то полезные для этого классификации. Надеюсь, что задача будет состоять, в основном, лишь в том, чтобы их найти и установить даты возникновения (жизни) соответствующих боле мелких таксонов (входящих в отрасли). Мне кажется, эти два этапа не так уж и сложно пройти, используя интернет (в том смысле, что это гораздо легче аналогичного пройденного этапа в биологии).
И следующий этап - это брать логарифм от количества упомянутых найденных более мелких таксонов и подбирать коэффициенты рассматриваемой степенно-логарифмической зависимости так, чтобы они наилучшим образом параметризовали динамику эволюции количества отраслей. Если окажется, что соответствие - удовлетворительно, то это будет означать, что рассматриваемая закономерность, действительно, является общеэволюционной. Т. е. свойственной для биологической и технологической эволюций. Что даст ещё один инструмент изучения техноэволюции, т. к. позволит, пользуясь ещё одной аналогией (качественной и количественной) с биоэволюцией, использовать наработанные в ней знания для изучения техноэволюции (как и наоборот).
******
Ну а теперь, подробнее, обосную существование рассмотренной закономерности применительно к биологии. А затем приведу косвенное свидетельство в пользу того, что эта закономерность работает и в технических системах.
Впервые степенно-логарифмическую зависимость в биологии, скорее всего, обнаружил в 2002 году. На рис. 1 приведены графики из его работы [1], которые иллюстрируют эту зависимость.
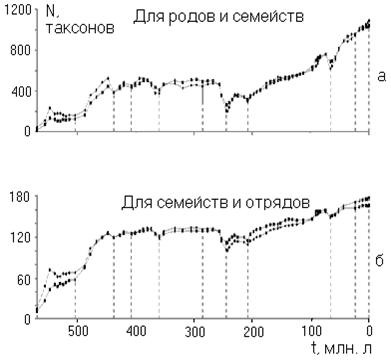 Рис. 1
Рис. 1
На рисунке "а" показана степенно-логарифмическая связь для количества морских родов и семейств (а = 0.2035, b= 4.00, коэффициент корреляции =0.98), на рисунке "б" – для морских семейств и отрядов (а = 1.310, b= 2.528, коэффициент корреляции также=0.98)[1]. По оси абсцисс показано время в миллионах лет до начала нашей эры.
Но если степенно-логарифмическая связь претендует на универсальность, то возникает проблема, связанная с неполнотой археологических данных. Действительно: чем глубже залегают археологические слои – тем меньше нам известно о существовавших, в соответствующие времена, биологических таксонах. И, как следствие, они могут быть ещё не все известны. Что ставит вопрос о том, можно ли пользоваться такими не полными данными для корректного, объективного поиска аналитической зависимости между количеством таксонов или нет?
Однако, если сравнить два графика зависимости от времени количества “существовавших”[2] таксонов, построенных по археологическим данным разных лет, то оказывается, что в целом, они, с точностью до константы, примерно, совпадают для всех биологических таксонов, кроме видов. Например, такая связь отражена на рис. 2. для семейств [2].

Рис. 2
На нижней части рисунка, показаны графики зависимости от времени количества морских семейств, известных по археологическим данным за 1982 и 1992 годы. На верхней части рисунка показано относительное процентное приращение количества этих семейств, между упомянутыми годами. По оси абсцисс показано время в миллионах лет до нашей эры, а так же, буквами, обозначены соответствующие геологические периоды.
Как видно – прирост, открытых за 10 лет семейств, составляет около 12 процентов. И, оказывается, что если бы этот прирост был даже десятикратным, то с точностью, примерно, плюс минус 20 процентов степенно-логарифмическая закономерность всё равно бы выполнялась, но только при другом коэффициенте пропорциональности (имеется в виду случай, когда количество семейств стоит в правой части (1)). Что связано с относительно медленным изменением логарифмической функции.
Т. е. проверка на наличие степенно-логарифмической закономерности мало чувствительна к, примерно, пропорциональному изменению количества таксонов, связанному с ростом полноты археологических данных. По этой же причине она мало чувствительна к тому, чтобы отдельно делать такую проверку только по археологическим данным для морских таксонов или одновременно для морских и сухопутных. Потому, что две последние величины, примерно, пропорциональны друг другу и имеют одинаковый порядок[3]. В связи с чем, в этой статье, я ограничусь анализом степенно-логарифмической закономерности лишь для морских организмов. Потому, что для морских организмов, в отличие от сухопутных, имеющаяся база данных позволяют вести анализ, начиная от родов и, кончая, типами (достаточно полные археологические данные по сухопутным родам сейчас отсутствуют). Но, при этом, буду иметь в виду, что закономерность претендует на всеобщность. Т. е. применительно к общему количеству таксонов. Т. е. морских и сухопутных вместе взятых.
Наличие низкой чувствительности степенно-логарифмической закономерности по отношению к неполноте данных всё же заставило меня усомниться в глубинном её смысле. Поэтому решил проверить закономерность по отношению к другим парам «соседних» таксонов, по которым имеются археологически данные, неполнотой которых, пожалуй, можно пренебречь. Это классы/отряды и типы/классы [3]. Однако, поначалу, был разочарован (рис. 3).
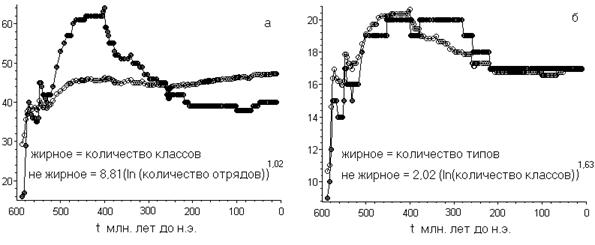 |
Рис. 3.
Степенно-логарифмическая зависимость за последние 580 миллионов лет между морскими отрядами и классами (рис. а, коэффициент корреляции равен 0,35), классами и типами (рис. б, коэффициент корреляции равен 0,87). Параметры подобраны методом наименьших квадратов путём минимизации дисперсии.
Как видно на рис. 3, особенно явное несоответствие со степенно-логарифмической закономерностью наблюдается для связи между классами и отрядами. Что является следствием того, что где-то 400 миллионов лет назад количество классов (так же и типов) начало сокращаться, в отличие от отрядов.
Но у меня возникла гипотеза, которая могла бы снять эту трудность. А именно, предположил, что сокращение количества классов (типов) происходило вследствие того, что характерное времена возникновения классов (типов) оказалось слишком великим по отношению к слишком выросшим темпам эволюции. Иначе говоря, предположил следующую картину.
Новые классы (типы) возникают из старых, но, вследствие ускоренного характера эволюции, старые классы (типы) ещё не успевают сложиться (возникнуть окончательно) прежде, чем возникают экологические ниши для возникновения новых классов (типов). В результате ещё не возникшие классы (типы), т. е. ещё в зародыше, начинают делиться, давая жизнь зародышам новых классов (типов), и т. д. Т. е., фактически, вследствие слишком выросшей (и постоянно растущей) скорости эволюции классы и типы перестали достаточно чётко оформляться по соответствующим (морфологическим, фенотипическим) признакам. Хотя по занимаемым экологическим нишам оформление, предположил, наступает. Иначе говоря, предположил, что классы (типы) прошли некую такую точку, после которой темпы эволюции биосферы выросли на столько, что сами условия (экологические ниши) для возникновения классов (типов), создаваемые самим процессом биологической эволюции, стали меняться столь стремительно, что новые классы (типы) в них просто перестали успевать возникать. Тогда как старые классы (типы) продолжили, постепенно, вымирать от «старости» (что свойственно для любых биологических таксонов [4]). В результате и начало происходить общее сокращение количества классов (типов).
Из сформулированного предположения следует, что если верна степенно-логарифмическая закономерность, то она выполнялась для пар классы/отряды и типы/классы до некоторого момента в прошлом. После которого, в качестве количества классов (типов), следует брать сумму количества классов (типов) с некоторым количеством экологических ниш, в которых эти классы (типы) ещё не успели явно оформиться.
И, оказалось, что, действительно, где-то до 380 миллионов лет назад степенно-логарифмическая закономерность для соотношений классы/отряды и типы/классы выполнялась (Рис. 4).
 |
Для этих, более крупных таксонов, неполнота археологических данных существенно уменьшается (её можно пренебречь). Поэтому для них логично ожидать более близкие друг к другу значения коэффициентов, нежели чем в выше рассмотренных случаях (Рис.1). Примерно это и наблюдается. Что так же иллюстрирует рис. 4.
Рис. 4
Степенно-логарифмическая зависимость между морскими отрядами и классами (рис. а, коэффициент корреляции равен 0,989),классами и типами (рис. б, коэффициент корреляции равен 0,989). Параметры подобраны методом наименьших квадратов путём минимизации дисперсии.
Таким образом, во всех рассмотренных случаях, в которых относительно хорошо выполняется степенно-логарифмическая закономерность, коэффициенты пропорциональности менялись, примерно, в пределах от 0,2 (роды/семейства) до 2,3 (семейства/отряды) со средним значением 0,77. А степени менялись, примерно, в пределах от 2,15 (классы/типы) до 4-х (роды/семейства) со средним значением 2,89. Т. е., если грубо, коэффициент пропорциональности имеет порядок единицы, а степень находится где-то около значения показателя экспоненты (который равен около 2,72).
Последние же два рассмотренных случая (Рис. 4) – особенно показательны. Потому, что они касаются столь крупных таксономических групп, что для них, за рассматриваемый период, скорее всего, можно пренебречь неполнотой данных. Например, количество всех типов и классов живых организмов сейчас уже почти полностью известно за последние, примерно, 500 млн. л. И, поэтому для них, соответствующие коэффициенты наиболее близки к реальным.
Эти коэффициенты, вероятно, можно так же уточнить, производя обобщение понятий тип и класс на случай, где-то после 380 миллионов лет назад. Когда образование новых типов и классов прекратилось, но, вероятно, продолжилось образование неких экологических ниш им соответствующих, в которых образование новых типов и классов, продолжилось бы, если бы не слишком сильно выросшие темпы эволюции. Выявление динамики роста количества этих ниш - потенциальных зародышей новых классов и типов, вероятно, и могло бы позволить уточнить соответствующие константы путём обобщения степенно-логарифмической закономерности для связи между классами и типами после 380 миллионов лет назад.
Обращу внимание на то, что выше рассмотренный разброс коэффициентов степенно-логарифмической зависимости может быть, во многом, связан с тем, что они обладают способностью очень сильно «плавать», не очень сильно меняя визуальное соотношение между точками и их параметризацией. Иначе говоря, от показателя степени, графики меняются не так уж и сильно. Например, если для зависимости из предыдущего графика (рис. 4б) показатель степени "руками" дорастить почти на единицу, оптимизируя при этом коэффициент пропорциональности (он уменьшается, примерно, в три раза), то изменение будет таким, как показано на рис. 5.
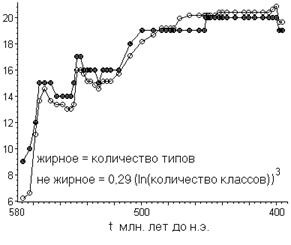 Рис. 5
Рис. 5
Степенно-логарифмическая зависимость между морскими классами и типами. Коэффициент корреляции 0,991. Показатель степени фиксирован «руками», а коэффициент пропорциональности оптимизировался путём минимизации дисперсии.
Рис. 5 иллюстрирует правдоподобность того, что коэффициенты рассматриваемой степенно-логарифмической зависимости для связей соседних таксонов, действительно, вполне могут быть, примерно, одними и теми же для всех соседних таксонов. И, вероятно, отличаются, в основном, вследствие либо случайных причин, либо вследствие неполноты данных (которая также может иметь случайную составляющую для разных интервалов времени), либо вследствие того, что рассматривается лишь морское биоразнообразие, а не совокупное. Причина небольшого отклонения от степенно-логарифмической зависимости между типами и классами также ещё состоит в относительной скачкообразности изменении количества типов. Поскольку типов не так много и их количество, со временем, меняется скачками, по сравнению с изменением количества классов. И поэтому изменение, со временем, количества типов просто не может вписаться в относительно плавные изменения для степени от логарифма классов.
Таким образом, коэффициент пропорциональности порядка единицы, а показатель степени около показателя экспоненты вполне подходят как претенденты на некоторую универсальность, которая будет более чётко выявлена по мере роста точности определения коэффициентов. Что наводит меня на мысль о существовании каких-то универсальных механизмов возникновения степенно-логарифмической зависимости, вероятно, связанных с фрактальной структурой биосферы, проявлением которой, вероятно, являются так же и ранговые распределения таксонов. Обнаружение же аналогичной, степенно-логарифмической, зависимости между какими-то технологическими таксонами позволило бы существенно повысить точность определения соответствующих коэффициентов. Поскольку для некоторых технологических таксонов неполноту данных, скорее всего, можно, практически полностью свести к нулю.
Упомянутое уточнение рассматриваемых коэффициентов позволило бы так же уточнить степенно-логарифмическую зависимость для биологических таксонов. А так же позволило бы реконструировать реальное количество биологических таксонов по количеству, известному из археологических данных. Что было бы существенным шагом в понимании эволюции биоразнообразия.
Упомянутую реконструкцию можно произвести, если иметь в виду, что количество известных, из археологических данных, таксонов, примерно, пропорционально реально существовавшему количеству. Действительно, если ![]() и
и ![]() – известное, из археологии, количество «соседних» таксонов, то
– известное, из археологии, количество «соседних» таксонов, то ![]() и
и ![]() – реально существовавшее. Где
– реально существовавшее. Где ![]() ,
, ![]() – некоторые константы, характеризующие неполноту данных. Тогда подставляя упомянутые реально существовавшие значения в степенно-логарифмическую зависимость (1) получаем:
– некоторые константы, характеризующие неполноту данных. Тогда подставляя упомянутые реально существовавшие значения в степенно-логарифмическую зависимость (1) получаем: ![]() . И, поскольку, считаем, что коэффициенты
. И, поскольку, считаем, что коэффициенты ![]() и
и ![]() известны из данных по технологическим таксонами, то отсюда, путём оптимизации по биологическим данным, можно, относительно точно, найти коэффициенты пересчёта
известны из данных по технологическим таксонами, то отсюда, путём оптимизации по биологическим данным, можно, относительно точно, найти коэффициенты пересчёта ![]() ,
, ![]() от количества известных, по археологическим данным, таксонов к реально существовавшим. Что существенно обогатило бы представление об эволюции биоразнообразия и позволило бы узнать вещи, которые, казалось бы, невозможно узнать никак иначе кроме как путём полной археологической реконструкции всех существовавших таксонов за рассматриваемый период (что чудовищно сложно). Точность поиска коэффициентов пересчёта можно ещё более улучшить, если вести оптимизацию по данным не одной какой-то пары «соседних» таксонов, а сразу по всем парам.
от количества известных, по археологическим данным, таксонов к реально существовавшим. Что существенно обогатило бы представление об эволюции биоразнообразия и позволило бы узнать вещи, которые, казалось бы, невозможно узнать никак иначе кроме как путём полной археологической реконструкции всех существовавших таксонов за рассматриваемый период (что чудовищно сложно). Точность поиска коэффициентов пересчёта можно ещё более улучшить, если вести оптимизацию по данным не одной какой-то пары «соседних» таксонов, а сразу по всем парам.
******
Но решение задачи обнаружения степенно-логарифмической зависимости по технологическим таксонам – не просто. Поэтому гипотезу о том, что технологические таксоны подчиняются той же степенно-логарифмической закономерности, что и биологические решил, предварительно, проверить косвенным образом. Используя, при этом, предположение о том, что количество некоторой категории технологических таксонов (пока, точно неизвестно какой) росло пропорционально народонаселению планеты, которое, примерно, подчиняется гиперболическому закону ~1/(2025 г.-t).
На основе знания ряда схожих процессов в социальной и биологической эволюциях у меня давно вертелось предположение о том, что, вследствие ускоренного характера эволюции, рост биологических видов, по своему ходу, постепенно перерос в рост технологических видов. Это предположение и использую для упомянутой косвенной проверки. А так же предположение о том, что народонаселение планеты, увеличиваясь по гиперболе, росло, примерно, пропорционально количеству неких технологических видов (вероятно, что пропорционально последнему растёт количество и видов человеческих изделий).
Иначе говоря, предполагаю, что, на некотором этапе биологической эволюции её темпы вырастают на столько, что новые экологические ниши начинают возникать столь быстро, что биологическая эволюция начинает не успевать давать «материал» для их заполнения. В результате, предполагаю, биологическая эволюция и начинает, постепенно сменяться технологической. Когда разные экологические ниши, вследствие недостаточной скорости биологической эволюции, начинают заниматься одним и тем же видом, который, для этого, начинает использовать орудия. Это может быть человек или предчеловек или даже какие-то другими виды, не являющиеся приматами и даже существующие параллельно с человеком по сей день. Например, вороны являются искусными изготовителями всяких палочек, крючочков и т. п., позволяющих им проделывать разные искусные ухищрения, чтобы доставать пищу, специальным образом спрятанную экспериментаторами.
Но для проверки сформулированной гипотезы нужно знать динамику роста количества всех биологических видов. А эту информацию существующие археологические данные дать не могут. Но когда я узнал про существование степенно-логарифмической зависимости между «соседними» биологическими таксонами, то возникла идея использования её (зависимость) для реконструкции динамики эволюции количества видов, используя то, что вид является «соседним» таксоном по отношению к роду. А динамика эволюции количества родов (морских организмов) относительно хорошо известна из археологических данных[4]. Иначе говоря, если верна гипотеза о том, что рост количества биологических видов был продолжен ростом количества неких технологических таксонов (эти таксоны называю технологическими видами), количество которых было пропорционально народонаселению, то натуральный логарифм от величены пропорциональной гиперболе народонаселения, возведённый в некоторую степень, должен описывать динамику количества биологических родов. Причём, соответствующие коэффициенты должны оказаться, примерно, такими же, как и выше полученные коэффициенты для степенно-логарифмической связи между соседними биологическими таксонами.
Проверка показала, что это предположение оказалось верным. Ниже изложу подробности. Но, прежде, хочу подробно остановиться на деталях обоснования предположения о том, что, действительно, эволюция биологических видов, постепенно, переросла в эволюцию видов технологической деятельности человека.
Для развития темы начну с рассмотрения парадокса. Для этого задамся вопросом о том, какие преимущества мог иметь человек, которые позволили ему держать эволюционное «первенство» перед своими биологическими конкурентами?
Ведь каждая из множеств специализаций (ниш), которую человек осваивал, «выходя» из дикой природы, в принципе, могла бы с более высоким успехом быть освоена более примитивными формами жизни. Например, ловить зайца лиса физически гораздо лучше приспособлена, чем человек. И это относится, практически, ко всем специализациям, которые осваивал человек «выходя» из природы. Тогда возникает вопрос, в чем было его преимущество?
Казалось бы, что первобытный человек, благодаря более высокому интеллекту, может использовать для поимки зайца гораздо более хитрую тактику, чем лиса. В результате чего в этом отношении он может оказаться успешнее лисы. Кроме того, человек может использовать приготавливаемые орудия, например, копье. Что увеличивает эффективность охоты. Однако представим себе такую фантастическую ситуацию, что у человека на генетическом уровне закрепился способ охоты на зайца с помощью каких то изготовляемых им орудий и стандартных, хитрых приёмов. Причем предположим, что способ изготовления орудий тоже закреплен на генетическом уровне (отдаленно похожая ситуация имеет место у некоторых муравьев, которые ведут «сельское» хозяйство, специально выращивая тлей, защищая их от опасностей, обеспечивая им укрытие и кормом и т. п.). В результате изготовление орудий на зайца и охота за ним у такого человека приняли бы инстинктивный характер. Закрепленные длительным отбором такие инстинкты стали бы гораздо более эффективными, нежели чем соответствующие приобретаемые навыки путем, преимущественно, обучения и практики (хотя, практика тоже нужна). Т. к. практика – относительно коротка. Поэтому рассматриваемому воображаемому человеку, по отношению к реальному, не нужен даже весь тот интеллект, который есть у реального человека, чтобы иметь над ним преимущество в охоте на зайца. Здесь ситуация полностью аналогична рассмотренной выше ситуации с зайцем и лисицей по отношению к человеку. Лисица имеет безусловное преимущество над человеком-одиночкой в поиске и поимке зайца, если человек не будет использовать орудия. Так и гипотетический человек, у которого поимка зайца и изготовление соответствующих орудий закреплены, благодаря длительному отбору, на инстинктивном уровне имеет преимущество (в плане поимки зайца) над обычным человеком. Тогда возникает вопрос, почему в реальности таких фантастических видов людей не возникало вследствие отбора (строго говоря, их нельзя было бы назвать людьми в силу предположения об относительной примитивности их интеллекта; назову таких людей людьми-зайцами)?
Можно предположить, что человека-зайца не возникло по той причине, что люди имеют гораздо более широкие специализации в добывании пищи и поэтому более гибки в поиске пропитания. Т. е., в противоположность рассматриваемому фантастическому человеку-зайцу, реальные люди могут охотиться и на других живых существ, например, оленей. Тогда как «человек-заяц», имея относительно ограниченные возможности по добыче пиши, был бы более уязвим по сравнению с настоящим человеком, имеющим более широкий круг специализаций. Однако тогда возникает вопрос, почему аналогично не возникли «люди-олени», «люди-мамонты» и т. д.? Такие «люди» все вместе взятые вытеснили бы настоящего человека во всех соответствующих узких специализациях. Т. к. они, каждый в своей специализации, имели бы несравненное преимущество над человеком. И тогда способность человека к широкому спектру специализаций не помогла бы ему выжить. И в результате становится не ясным, почему вообще возникли люди? В чем и состоит парадокс. Который ставит вопрос о том, что мешало возникновению рассмотренных фантастических людей с рассмотренными относительно узкими специализациями?
В процессе эволюции у разных живых существ появляются разные биологические приспособления. Благодаря этому они открывают для себя новые экологические ниши. Которые заполняются всё более быстро растущим количеством видов (ускоренность темпов эволюции). Для наглядности предположу, что этот рост – гиперболический (но для дальнейшего изложения достаточно предположения об его ускоренном характере): 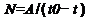 . Виды, заполняющие ниши, взаимодействуя с другими видами, создают основу для возникновения других экологических ниш, которые опять заполняются новыми видами, тем самым, создавая основу для дальнейшего, всё более ускоренного роста количества ниш других новых видов и т. д. В результате получается, что чем больше видов – тем выше прирост соответствующих новых экологических ниш, занимаемых новыми видами за единицу времени. Этот прирост равен
. Виды, заполняющие ниши, взаимодействуя с другими видами, создают основу для возникновения других экологических ниш, которые опять заполняются новыми видами, тем самым, создавая основу для дальнейшего, всё более ускоренного роста количества ниш других новых видов и т. д. В результате получается, что чем больше видов – тем выше прирост соответствующих новых экологических ниш, занимаемых новыми видами за единицу времени. Этот прирост равен 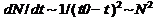 . Новые виды возникают из старых путем биологической эволюции старых. Среднее количество видов, возникающее за характерное время жизни вида из одного старого вида, равно произведению скорости прироста видов
. Новые виды возникают из старых путем биологической эволюции старых. Среднее количество видов, возникающее за характерное время жизни вида из одного старого вида, равно произведению скорости прироста видов 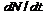 на характерное время жизни вида T и всё деленное на общее количество видов
на характерное время жизни вида T и всё деленное на общее количество видов ![]() :
: ![]() . Или, в силу гиперболического закона роста
. Или, в силу гиперболического закона роста ![]() , эта величина должна расти пропорционально произведению количества видов на их характерное время жизни:
, эта величина должна расти пропорционально произведению количества видов на их характерное время жизни: ![]() . Как видно - эта величина, со временем должна неограниченно расти в силу гиперболического роста количества видов, если предположить, что характерное время жизни вида не убывает быстрее роста гиперболы. Прямых, надёжных, усреднённых археологических данных по времени жизни видов не существует. Однако, по всей видимости, характерное время жизни биологических таксонов, начиная от родов и выше, в процессе эволюции, не убывает. Например, для морских родов это хорошо известно по работе [5], где приводятся данные о линейном росте средней продолжительности существования морских родов в течение фанерозоя (примерно, последние 600 миллионов лет). И, скорее всего, в глобальном плане, средняя продолжительность существования и для видов тоже не сокращалась. А, поэтому, в силу сказанного, среднее количество видов, «рождённых» одним видом за характерное время существования вида (время жизни вида) должно расти.
. Как видно - эта величина, со временем должна неограниченно расти в силу гиперболического роста количества видов, если предположить, что характерное время жизни вида не убывает быстрее роста гиперболы. Прямых, надёжных, усреднённых археологических данных по времени жизни видов не существует. Однако, по всей видимости, характерное время жизни биологических таксонов, начиная от родов и выше, в процессе эволюции, не убывает. Например, для морских родов это хорошо известно по работе [5], где приводятся данные о линейном росте средней продолжительности существования морских родов в течение фанерозоя (примерно, последние 600 миллионов лет). И, скорее всего, в глобальном плане, средняя продолжительность существования и для видов тоже не сокращалась. А, поэтому, в силу сказанного, среднее количество видов, «рождённых» одним видом за характерное время существования вида (время жизни вида) должно расти.
С другой стороны эта величина не может превышать некоторого предела в силу биологических ограничений на скорость «ветвления» видов. Эти ограничения связаны с конечностью скорости генетических модификаций, которые могут приводить к возникновению новых видов. Скажем, в естественных условиях, один вид вороны за сто лет не в состоянии дать сто видов других ворон. Потому, что за это время сменится не столь много поколений ворон, в течение которых успеют оформиться различные биологические изменения, соответствующие разным видам (заполняющим соответствующие биологические ниши). Таким образом, рано или поздно, ускоренный рост количества видов (в рассматриваемом примере - гиперболический) не сможет выполняться в силу биологических ограничений на способность каждого из видов за время своего существования порождать новые виды, которые бы своим количеством поддерживали ускоренный (гиперболический) закон роста количества видов. Возникновение такой ситуации означало бы начало глобального завершения «естественного» процесса роста (т. е. происходящего благодаря естественному отбору) биологического видообразования. Эта ситуация полностью аналогична ситуации с гиперболическим ростом народонаселения. Которое не может следовать гиперболе неограниченно долго вследствие того, что за время жизни человечество не может обеспечить воспроизведение потомства выше некоторого предела, связанного с биологическим ограничением по скорости деторождения женщиной.
Примером непоспевания заполнения ниш может быть заполнение ниш, связанное с изменением физиологии строения, способствующее более эффективной добыче пищи того или иного рода. Например, рассмотрим рост хобота слона. Зададимся вопросом, что будет, когда новые ниши за единицу времени начинают возникать в столь большем количестве, что, образно говоря, существа с соответствующими этим нишам новыми «хоботами» не успеют возникать путем эволюционного видоизменениям уже существующих существ?
Предполагаю, что тут, образно говоря, человек и берет в руки палку. Т. е. возникают существа, которые приобретают способность «искусственно» «доращивать» себе «органы», в замен тех не возникших существ которые не успели появиться благодаря слишком медленной для этого биологической эволюции. Откуда, предполагаю, и возникает эволюционная потребность и необходимость роста интеллекта. Т. к. используя интеллект, но, особо, не меняя своей физиологии, эти существа приобретают способность занимать те ниши, которые продолжают возникать со все увеличивающейся скоростью. Поэтому рассмотренные выше люди-зайцы, люди-олени и т. п. просто не успели возникнуть. И поэтому они и не составили конкуренции человеку. Благодаря чему и разрешается рассмотренный выше парадокс. А если бы подобные люди и начали возникать, то они не смогли бы вытеснить человека потому, что прежде чем они бы возникли – появилось бы (вследствие «большой» и постоянно увеличивающейся, в глобальном плане, скорости нишеобразования) «огромное» количество новых ниш, которые бы человек занять смог, используя «высокую» скорость эволюции своих приспособлений, а другие существа занять бы их (ниши) не успели. И тем самым, в дальнейшем, человек всегда бы «сильно» опережал в приспосабливаемости из-за своей технологической мобильности.
Этот механизм вполне соответствует эмпирической закономерности, отмечаемой [5] для роста, в процессе эволюции морских животных, эврибионтности видов. Состоящей в том, что вид в процессе эволюции, в среднем, занимает все боле широкую биологическую нишу. Т. е. в рамках одной и той же биологической организации виды, в целом, осваивают все большее количество источников пропитания. Пределом чего явилось появление человека, который, с помощью приспособлений, в принципе, способен занимать практически любые биологические ниши, оставаясь в рамках одной и той же биологической организации.
В пользу верности выше описанного представления о причине возникновения технологической эволюции (человека) свидетельствует также процесс «глобализации» мировой экономики. Что связано с тем, что в процессе социальной эволюции закон роста эврибионтности должен принять форму «сближения» между разными культурными и национальными общностями.
Действительно, несмотря на то, что в «момент» возникновения человека эволюционный рост разнообразия занимаемых им потенциальных видовых ниш стал происходить, в основном, в рамках одного биологического вида – в рамках разных этнических и культурных вариаций он (рост) все-таки сразу прекратиться не мог. Иначе говоря, в «момент» возникновения человека этнические, культурные вариации представляли собой ни что иное, как «зачатки» будущих биологических видов, которые возникли бы, если бы скорость прироста новых видов человеческой деятельности не была бы слишком «большей». И такие виды (людей), действительно, некоторое время возникали. Например, «классические» неандертальцы (вид человека), специализировались, преимущественно, на охоте на мамонтов в северных широтах. Но рано или поздно, вследствие ускоренного характера эволюции, рассматриваемая скорость прироста (теперь уже в процессе социальной эволюции) не может не стать столь высокой, что и культурные различия между людьми должны стереться. Стереться как «стерлись» биологические различия между предками людей – гоминидами («стирание» происходит путем конкурентного вытеснения менее универсальных в своих приспособлениях, поведении и т. п. более универсальными). И эта тенденция, действительно, прослеживается в современном мире (глобализация).
Таким образом, рассмотрение причины появления человека как следствие ускоренного характера биологической эволюции позволяет найти подход в поиске преемственности между социальной и биологической эволюциями.
В частности, в соответствие со сказанным, в будущем Землю ждет одна (единая) человеческая культура, один язык, которые, в силу сказанного, будут наиболее приспособлены к «очень» быстрой социальной эволюции.
Из сказанного следует, что один из аспектов единства социальной и биологической эволюций состоит в том, что в процессе биологической эволюции биологические таксоны должны «плавно» переходить в некие социальные (технологические) аналоги, постепенно принимая, полную эволюционную самостоятельность от биологической основы. Т. к. движущие принципы роста новых ниш не отменяются тем фактом, что они теряют способность расти в рамках соответствующих разных биологических организациях. В соответствии со сказанным, с возникновением человека таксоны должны, постепенно, начать принимать форму не биологических специализаций, а технологических. С функциональной точки зрения причин гиперболического роста нет разницы между способом реализаций специализаций. Не важно: растет ли количество ниш благодаря росту количества биологических специализаций, либо социально-технологических. В том и другом случае должны наблюдаться некие общие для биологической и социальной эволюций процессы. Которые являются только функцией количества неких эволюционных единиц (биологических эволюционных единиц, которыми могут быть, например, биологические виды или продолжающие их социальные аналоги). Что позволяет поставить задачу поиска сходства в развитии неких биологических образований с некими социальными, которые должны явиться их эволюционными продолжениями. А это, вероятно, откроет путь решения ряда теоретических вопросов сугубо социальной эволюции на основе известных решений задач-аналогов из биологической эволюции и наоборот.
А в пользу того, что количество технологических видов росло пропорционально народонаселению говорит то, что технологические виды – это, по сути, виды человеческих специализаций, т. е. специальностей (от которых логично ожидать, что их количество росло пропорционально народонаселению, а так же логично ожидать, что пропорционально им росло количество видов человеческих изделий). Потому, что, по своему определению, технологические виды являются продолжением развития биологических специализаций, поначалу, реализовавшихся в рамках разных биологических видов. Высокие темпы эволюции привели лишь к тому, что они (специализации) стали реализовываться в рамках одного вида. Тем самым, постепенно, приобретя эволюционную самостоятельность, независимость от биологической эволюции.
Развивая эту мысль, можно ожидать, что не только биологические виды имеют своих эволюционно-функциональных последователей в технологической эволюции. Но и биологические роды, семейства, классы, типы и т. п. Что и явилось у меня одной из основ для предположения о том, что и в процессе технологической эволюции выполняется степенно-логарифмическая связь между соответствующими «соседними» технологическими таксонами. Потому, что, в силу выше сформулированной гипотезы, они являются теми же биологическими таксонами, только, в силу слишком выросших темпов эволюции, принявшими технологическую форму.
В этой связи наличие гиперболических ранговых распределений в техноценозах и биоценозах становится тоже более понятным. Потому, что, получается, что первые являются просто ни чем иным как формой вторых, вынужденно возникшей из-за слишком выросших темпов эволюции. В результате чего эволюция биологических таксоны начала постепенно функционально замещаться эволюцией технологических, а ранговые распределения остались.
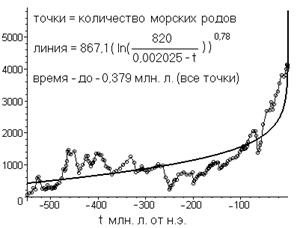 Чтобы не быть голословным, теперь приведу обещанные подробности. Это будет график, связывающий гиперболу народонаселения ~1/(0,002025 мл. л.-t) и динамику количества родов. Но, прежде, ещё замечу, что, если верно выше сказанное, то на промежуточном этапе между биологической и социальной эволюциями степенно-логарифмическая связь между гиперболой народонаселения и количеством родов должна нарушаться. Т. к. эта связь должна, в силу сказанного, выполняться лишь для суммы количества биологических видов и количества эволюционно продолжающих их технологических видов. Эта ситуация вполне аналогична выше описанной ситуации с классами и типами, когда начало происходить замедление их роста. Поэтому на промежуточном этапе от неё (от степенно-логарифмической связи) можно ожидать нарушения. И это действительно, видно по рис. 6 [3].
Чтобы не быть голословным, теперь приведу обещанные подробности. Это будет график, связывающий гиперболу народонаселения ~1/(0,002025 мл. л.-t) и динамику количества родов. Но, прежде, ещё замечу, что, если верно выше сказанное, то на промежуточном этапе между биологической и социальной эволюциями степенно-логарифмическая связь между гиперболой народонаселения и количеством родов должна нарушаться. Т. к. эта связь должна, в силу сказанного, выполняться лишь для суммы количества биологических видов и количества эволюционно продолжающих их технологических видов. Эта ситуация вполне аналогична выше описанной ситуации с классами и типами, когда начало происходить замедление их роста. Поэтому на промежуточном этапе от неё (от степенно-логарифмической связи) можно ожидать нарушения. И это действительно, видно по рис. 6 [3].
Рис. 6.
По оси абсцисс отложено время, отсчитываемое от нашей эры (до неё время считается отрицательным; на предыдущих графиках оно считалось положительным). Параметры линии подобраны минимизацией дисперсии.
Как видно, полученные параметры очень сильно отличаются от характерных параметров степенно-логарифмической связи между соседними биологическими таксонами. Например, коэффициент пропорциональности оказался аж на три порядка больше. Однако оказывается, что если исключить из оптимизации шесть самых правых точек, то график и параметры оказываются уже приемлемыми (рис. 7).
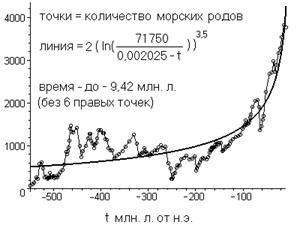 Рис 7
Рис 7
Как и выше, по оси абсцисс отложено время, отсчитываемое от нашей. Оптимизировался (минимизацией дисперсии) только один параметр – коэффициент пропорциональности гиперболы народонаселения (значение 71750). Остальные параметры подобраны «на глаз» из соображения близости к аналогичным коэффициентам для связи между соседними биологическими таксонами.
Как видно, параметры степенно-логарифмической связи вполне вписываются в интервал, в котором лежат аналогичные параметры, связывающие «соседние» биологические таксоны. А точки, которые были выкинуты, примерно, соответствуют пику расцвета человекообразных обезьян. Если не выкидывать эти точки, то вписаться в упомянутый интервал (при приемлемом соответствии точек и графика) не получится не при каких параметрах. С расцветом же человекообразных обезьян, хронологически, эволюция биосферы, вероятно, вступила в новый качественный этап роста, о котором говорилось выше. Т. е. когда часть вновь возникающих экологических ниш, которые ранее заполнялись, в основном, только вновь возникающими видами, стали, заполняться какими-то одними видами (не обязательно человекообразными обезьянами одного вида и не обязательно обезьянам).
Заключение.
Подведу некоторые итоги.
Если подтвердится наличие рассмотренной степенно-логарифмической закономерности не только в биологии, но и в технике, то это будет означать новый критерий для построения численных фундаментальных моделей эволюции. Как социальной, так и биологической. Что очень важно в том смысле, что это позволит переносить знания, накопленные в биологии на социальный случай и наоборот. А так же важно в том смысле, что таких критериев, скорее всего, не так уж много. Мне известен только один (работающий как для биологической, так и для социальной или технологической эволюций) – это гиперболические, ранговые распределения (и их модификации). Использование двух критериев, вероятно, сможет существенно снизить круг поиска среди разных моделей эволюции.
Кроме того, подтверждение наличия степенно-логарифмической закономерности в технике будет означать более точное знание её коэффициентов. Т. к. для технических систем неполнота данных гораздо меньше, чем в археологии. Используя эти, более точные, коэффициенты в биологии можно реконструировать реальную динамику роста биоразнообразия планеты на протяжении, примерно, последних 500 миллионов лет исходя из далеко не полных археологических данных. Более точное знание рассматриваемых коэффициентов позволит так же значительно более точно реконструировать динамику роста количества видов по количеству родов. А, вместе с этим, значительно более точно подтвердить пропорциональность динамики роста количества видов гиперболе народонаселения планеты. Если эта пропорциональность подтвердится с ещё более высокой точностью, то это будет серьёзным аргументом в пользу выше описанной гипотезы возникновения человека как следствие ускоренного характера биологической эволюции. Когда темпы эволюции вырастают на столько, что заполнение всех вновь возникающих экологических ниш становится невозможным только отдельными видами из-за ограниченной скорости биологической эволюции. И это заполнение, всё в большей степени, начинает происходить одним видом с использованием орудий и приспособлений.
Верность этой гипотезы будет означать нахождение глубинной, «генетической» связи между процессами социальной и биологической эволюций. Что позволит переносить огромные накопленные эволюционные знания из социально-технологической области на биологическую и наоборот. Кроме того, появится ещё один, третий, численный критерий, связывающий социальную и биологическую эволюции – это гиперболический рост (гипербола народонаселения планеты, продолжающая гиперболический рост количества видов).
Совместное использование упомянутых критериев, вероятно, может значительно облегчить задачу математического моделирование эволюции и более глубоко понять её механизмы. В частности, вероятно, позволит привлечь представление о фрактальности эволюции и математически его вписать в выше упомянутые численные закономерности.
Литература.
[1] “Проблемы эволюции таксономического разнообразия”, 2002, http://macroevolution. *****/dmitrie1.htm
[2] J. John Sepkoski, Jr, Paleobiology, Vol. 19, No.1. (Winter, 1993), pp. 43-51, “Ten Years in the Library: New Data Confirm Paleontological Patterns”.
[3] Соответствующие данные по динамике эволюции количества рассматриваемых таксонов любезно выслал . Они были получены на основе данных Дж. Сепкоски. Последние можно скачать с сайта http://macroevolution. *****
[4] Кратко ознакомиться с общим представлением об этом процессе можно в сетевой публикации «Этапность в эволюции таксонов (филогенетический цикл)», http://macroevolution. *****/cycle. htm
[5] , «О механизмах роста таксономического разнообразия морской биоты в фанерозое», Палеонтологический журнал, 2002, № 2, с. 3-13, http://macroevolution. *****/meanps. htm .
[1] Квадратиками показаны фактические значения, крестиками - вычисленные. Что, в показанном масштабе, не заметно. Чтобы увидеть различие в обозначениях точек лучше всего рассмотреть графики из статьи по ссылке в увеличенном виде.
[2] если бы археологически данные были бы столь полны, что были бы известны даты жизни всех, когда либо существовавших, в течение рассматриваемого периода времени, таксонов, то кавычки можно было бы убрать
[3] Один и тот же таксон может иметь как морских, так и сухопутных представителей. Таксон считается морским (или сухопутным), если он имеет хоть одного морского (сухопутного) представителя. В случае таксона тип морские и сухопутные таксоны, совпадают друг с другом. В случае классов – совпадение, практически, полное. Один и тот же таксон может иметь как морских, так и сухопутных представителей. Таксон считается морским (или сухопутным), если он имеет хоть одного морского (сухопутного) представителя. В случае таксонов тип и класс морские и совокупные таксоны (морские вместе с сухопутными), примерно, совпадают друг с другом.
.
[4] Напомню, что корректнее было бы использовать суммарное количество морских и сухопутных родов (достаточно полные данные по последним сейчас отсутствуют), но это не является принципиально важным. Т. к. морское и сухопутное биоразнообразия, в процессе эволюции, растут, примерно, пропорционально друг другу и имеют, примерно, одинаковый порядок. А степенно-логарифмическая связь, в силу присутствия логарифма, крайне низко чувствительна к, примерно, пропорциональному изменению количества таксонов даже в разы.
[5] Личное сообщение.
