
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
на правах рукописи
Морфофункциональная оценка роли P2Y-рецепторов в пищеводе при ахалазии
03.03.04 – клеточная биология, цитология, гистология
03.03.01 - физиология
Автореферат
диссертации на соискание ученой степени
кандидата медицинских наук
Казань – 2010
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность работы
Изучение роли метаботропных P2Y-рецепторов в регуляции физиологических функций организма является актуальной проблемой современной медицины (, , 2009). Экспрессия P2Y-рецепторов выявлена на плазматической мембране практически каждой клетки организма млекопитающих, и они участвуют во всех ключевых клеточных процессах (, 2005; Burnstock G., 2009).
В настоящее время активно исследуют участие различных типов P2Y-рецепторов в регуляции сократительной активности мышц. В экспериментах на препаратах ушка правого предсердия человека активация P2Y-рецепторов вызывает снижение амплитуды сокращений, а их блокирование – обратный эффект (, 2007). Показано, что активация P2Y-рецепторов пиримидиновым нуклеотидом УТФ вызывает расслабление гладкомышечных клеток пещеристого тела (Calvert R.C., 2008) и толстой кишки (Van Crombruggen К. et al., 2007).
Необходимо отметить, что отсутствуют сведения о роли P2Y-рецепторов в регуляции сократительной активности ГМК пищевода. Вместе с тем, модуляция работы P2Y-рецепторов представляется перспективным подходом в коррекции нарушений моторики пищевода, наблюдающихся при ахалазии – одного из наиболее часто встречающихся заболеваний пищевода. Среди всех функциональных нарушений пищевода ахалазия составляет свыше 31% (Belsei R., 1966, , 2000). Этиология заболевания неизвестна (Dughera L. et al., 2008), в то же время выдвигаются следующие теории развития ахалазии пищевода (АП): теория психических нарушений (, 1977), генетическая (Stein D.T., Knauer C.M., 1982., Frieling T. et al., 1988., Sachdev A. et al., 2004) и вирусная (Jones D.B. et al., 1983, Robertson C.S., Martin B.A., Atkinson M., 1993, Castagliuolo I. et al., 2004).
В основе патогенеза лежит повышение тонуса нижнего пищеводного сфинктера (НПС) и нарушение перистальтики пищевода вследствие дегенерации тормозных нейронов межмышечного (ауэрбаховского) нервного сплетения, которые отвечают за релаксацию гладкой мускулатуры НПС во время акта глотания (Park W., Vaezi M. F., 2005). В результате этого нарушается прохождение пищи из пищевода в желудок, что приводит к дилатации пищевода и кахексии пациента (Farrokhi F., Vaezi MF., 2009). Опасность этого заболевания заключается в высоком риске развития плоскоклеточного рака и аденокарциномы пищевода на фоне ахалазии (Zendehdel К. et al., 2007). Несмотря на широкий спектр используемых методов лечения АП, результаты часто неэффективны и имеют ряд серьезных осложнений (Bruley des Varannes S., Scarpignato C., 2001., Iqbal A. et al., 2006).
Отсутствие данных об экспрессии P2Y-рецепторов в стенке пищевода, о роли пуринергических механизмов в регуляции сократительной функции пищевода в норме и при патологии пищевода делает данное исследование актуальным. Кроме того, ответ на поставленные вопросы позволит в дальнейшем значительно продвинуться в понимании роли P2Y-рецепторов в регуляции физиологических функций и развитии заболеваний желудочно-кишечного тракта, открыть новые возможности в медикаментозной терапии при помощи препаратов с пуринергическим механизмом действия.
Цель и задачи исследования
Целью настоящего исследования явилось изучение экспрессии различных типов P2Y-рецепторов в нижней трети пищевода и их роли в регуляции сократительной активности пищевода в норме и при ахалазии.
В соответствии с этой целью были поставлены следующие конкретные задачи:
1. Провести сравнительное исследование экспрессии P2Y1, P2Y2 и P2Y4-рецепторов в нижней трети пищевода человека в норме и при ахалазии.
2. Выполнить морфофункциональный анализ результатов, полученных на экспериментальной модели ахалазии у крыс и провести оценку соответствия клинической и морфологической картине заболевания у человека.
3. Провести сравнительное исследование сократительной активности ГМК нижней трети пищевода и НПС крыс в норме и на фоне экспериментальной ахалазии.
4. Провести сравнительное исследование экспрессии P2Y1, P2Y2 и P2Y4-рецепторов в нижней трети пищевода крысы в норме и при экспериментальной ахалазии.
5. Оценить изменения сократительных ответов ГМК нижней трети пищевода и НПС крыс под действием различных агонистов и антагонистов P2Y-рецепторов в норме и при экспериментальной ахалазии.
Научная новизна
В диссертационной работе впервые проведено комплексное морфофизиологическое изучение роли P2Y-рецепторов в регуляции сократительной активности пищевода. Было установлено, что в норме у человека и крысы P2Y1,2,4-рецепторы локализуются во всех слоях стенки нижней трети пищевода. Впервые был проведен детальный морфофункциональный анализ течения экспериментальной ахалазии (ЭА) у крысы и доказана адекватность данной модели клинической и морфологической картине заболевания человека. Дана детальная характеристика нарушений сократительной активности нижней трети пищевода и НПС in vitro при ЭА у крыс. Также показано, что блокаторы P2Y-рецепторов увеличивают амплитуду медленных сократительных ответов НПС, тогда как активаторы не оказывают влияния. При ЭА эффекты как агонистов, так и антагонистов P2Y-рецепторов значительно видоизменяются, что свидетельствует о выраженном нарушении метаботропной пуринергической регуляции НПС.
Научно-практическая ценность
Выполненная работа расширяет знания о пуринергических механизмах регуляции функции пищевода и детально характеризует участие различных типов P2Y-рецепторов в сократительной активности нижней трети пищевода. В ходе исследования была разработана модифицированная методика ЭА у крыс, выполнен морфофункциональный анализ полученных результатов и доказана адекватность модели ЭА клинической и патологоанатомической картине заболевания у человека. Таким образом, нами было показано, что в дальнейшем данная модель может быть использована для разработки и изучения современных методов лечения ахалазии пищевода. Полученные экспериментальные данные значительно расширяют и дополняют наши представления об участии пуринорецепторов в регуляции функций пищеварительного тракта в норме и патологии и являются перспективными в плане разработки новых подходов к лечению ахалазии при помощи лекарственных препаратов с пуринергическим механизмом действия.
Основные положения диссертации, выносимые на защиту:
1. P2Y1,2,4-рецепторы локализуются в стенке пищевода человека и крысы в норме; при ахалазии у человека и при экспериментальной ахалазии у крысы происходит изменение экспрессии вышеуказанных типов P2Y-рецепторов.
2. Активность P2Y-рецепторов оказывает угнетающее влияние на сократительную активность ГМК нижнего пищеводного сфинктера крысы в норме. При экспериментальной ахалазии у крыс происходит выраженное нарушение метаботропной пуринергической регуляции нижнего пищеводного сфинктера.
Личный вклад диссертанта
Приведенные в работе данные получены при личном участии соискателя на всех этапах работы, включая составление плана исследования, постановку задач, выбор методов исследования, проведение экспериментов, обработку экспериментальных данных и оформление публикаций. Соискателем впервые в России методом иммуногистохимии с использованием LSAB-kit (DAKO) изучена экспрессия P2Y1,2,4-рецепторов в нижней трети пищевода человека и крысы в норме и при ахалазии, дана детальная характеристика нарушений сократительной активности пищевода при ЭА.
Достоверность полученных данных
Достоверность полученных данных достигнута использованием достаточного количества образцов для исследования, конкретной постановкой и решением поставленных задач с использованием статистического метода. Достоверность различий между результатами оценивали по t-критерию Стьюдента.
Сведения об апробации результатов диссертации
Основные результаты диссертационной работы доложены на следующих конференциях и форумах: 12 Международной Пущинской школе-конференции молодых ученых «Биология-наука XXI века» (Пущино, 2008), Всероссийской конференции «Научное наследие академика . Структурные и функциональные основы эволюции функций, физиология экстремальных состояний» (Санкт-Петербург, 2008), XIV Всероссийской научно-практической конференции «Молодые ученые в медицине» (Казань, 2009), XV межгородской конференции молодых ученых «Актуальные проблемы патофизиологии» (Санкт-Петербург, 2009), XV Всероссийской научно-практической конференции «Молодые ученые в медицине» (Казань, 2010), совместном заседании кафедр гистологии, цитологии, эмбриологии и нормальной физиологии ГОУ ВПО «КГМУ Федерального агентства по здравоохранению и социальному развитию».
Публикации материалов исследования
По материалам диссертации опубликовано 10 научных работ, в том числе 2 статьи в ведущих научных рецензируемых журналах, рекомендованных Высшей аттестационной комиссией для опубликования основных результатов диссертации. Общий объем публикаций - 1.8 у. п. л., в т. ч. авторский вклад – 1.1 у. п. л..
Внедрение результатов работы
Полученные данные используются при чтении лекций на кафедрах гистологии, цитологии, эмбриологии и нормальной физиологии Казанского государственного медицинского университета. Кроме того, результаты исследования внедрены в лечебный процесс Республиканского Клинического Онкологического Диспансера Министерства Здравоохранения Республики Татарстан (РКОД МЗ РТ).
Структура и объем диссертации
Диссертация объемом 111 страниц состоит из введения, обзора литературы, описания методики исследования, результатов исследования и их обсуждения, заключения, выводов и списка цитируемой литературы. Список цитируемой литературы включает 145 названий, из них 25 отечественных и 120 иностранных авторов. Диссертация содержит 34 рисунка и 3 таблицы.
СОДЕРЖАНИЕ РАБОТЫ
Объекты и методы исследования
Опыты проводили на 310 беспородных белых крысах массой 180–200 г. Животные содержались в стандартных условиях вивария в пластмассовых клетках с наполнителем при температуре 18–20оС, имели свободный доступ к пище и воде.
Для морфологического исследования также использовали образцы тканей человека:
1) биоптаты мышечной стенки нижней трети пищевода на 1-2 см выше кардиального сфинктера 35 пациентов, страдавших ахалазией пищевода 3 степени с длительностью заболевания от 9 месяцев до 13 лет, взятые во время операции лапароскопической кардиомиотомии с фундопликацией по Геллеру/Дору во втором торакальном отделении РКОД МЗ РТ (опытная группа).
2) кусочки стенки пищевода 34 пациентов второго торакального отделения РКОД МЗ РТ, больных раком средней/нижней трети пищевода, не получавших предоперационную лучевую терапию. Пациентам выполнили операцию трансхиатальную экстирпацию пищевода, после которой макропрепарат незамедлительно направлялся на плановое гистологическое исследование в патологоанатомическое отделение РКОД МЗ РТ, во время которого вырезали кусочки здоровой стенки пищевода на протяжении всей толщи стенки размером 1х1см на значительном расстоянии от опухоли (контрольная группа).
У всех испытуемых после проведения разъяснительной беседы было получено письменное добровольное согласие на участие в исследовании, которое проведено с разрешения Этического Комитета Республики Татарстан (выписка из протокола Бюро №1 от 01.01.01г.).
Моделирование ахалазии пищевода на крысах
Исходя из данных зарубежной литературы первая серия крыс была прооперирована согласно стандартной, опубликованной ранее методике (Goto S., Grosfeld J. L., 1989, Febronio L.H. et al., 1997). Учитывая высокую послеоперационную летальность, в дальнейшем животные были подвергнуты модифицированной операции экспериментальной ахалазии. Все хирургические манипуляции с животными проводили под уретановым наркозом (600 мг/кг внутрибрюшинно). В асептических условиях выполняли верхнюю срединную лапаротомию и осуществляли доступ к брюшной части пищевода. Под брюшную часть пищевода подкладывали протектор (отрезок из плотного полиэтилена 4х1.5 см.), затем осуществляли аппликацию на выделенную часть пищевода марлевой турунды, смоченной 0,2% раствором бензалкония хлорида – (БХ), (Sigma, США) в течение 30 минут. Необходимо отметить, что БХ является синтетическим анионовым детергентом, обладающим неспецифическим механизмом действия, приводящим к гибели нейронов (Primor N., 1986).
За этот промежуток времени турунду дважды по мере высыхания капельно смачивали БХ из инсулинового шприца. Затем брюшную полость зашивали непрерывным обвивным швом. Контрольную группу («ложно-оперированную») составили животные, которым при прочих одинаковых условиях раствор БХ был заменен на физиологический раствор. Интактную группу составили животные, которым не проводилась хирургическая операция. Для анализа течения ЭА производился контроль массы животных при помощи ежедневного взвешивания на лабораторных весах (МЕРА ВМ-2/6, Россия), начиная со дня операции и до 20-х суток послеоперационного периода. На 21-е сутки после операции крысам под уретановым наркозом 600мг/кг выполнили верхнюю срединную лапаротомию и измерили давление в нижней трети пищевода и НПС с помощью двухмиллиметрового датчика, состоящего из эластического резинового шарика и эпидурального катетера, который соединен с манометром.
Гистологические и иммуногистохимические исследования
Материал фиксировали в 10% нейтральном забуференном формалине, обезвоживали и заключали в парафин по стандартной методике. Срезы с парафиновых блоков выполняли серийно на ротационном микротоме Leica RM 2125 RT, (Германия), толщиной 3-5 мкм и окрашивали гематоксилином и эозином. Для иммуногистохимического анализа по локализации P2Y1,2,4–рецепторов, а также белков S100 и виментина использовали моноклональные антитела (разведение 1:100, Sigma) (табл. 1) и LSAB-kit (DAKO).
Таблица 1
Антитела, использованные в иммуногистохимическом исследовании
Антитело | Производитель | Разведение |
P2Y1-рецептор | Sigma (США) | 1:100 |
P2Y2-рецептор | Sigma (США) | 1:100 |
P2Y4-рецептор | Sigma (США) | 1:100 |
Белок S100 | DAKO (Дания) | 1:100 |
Виментин | DAKO (Дания) | 1:80 |
Миографическое исследование
Эксперименты по исследованию сократимости пищевода проводили на установке Power Lab, оборудованной датчиками силы MLT 050/D (AD Instruments, США). У крыс под наркозом иссекали нижнюю треть пищевода вместе с кардиальной частью желудка. Далее из области НПС выделяли циркулярную полоску, а из нижней трети пищевода – продольную полоску длиной 7-8 мм и диаметром 2-4 мм. В течение препаровки и всего эксперимента препараты пищевода находились в перфузионном растворе следующего состава (в мМ): NaCl -125, KCl - 2.5, CaCl2 –2, MgSO4 – 1, NaH2PO4 – 1.25, NaHCO3 – 25, глюкоза - 11. Перед экспериментом раствор аэрировали карбогеном в течение часа. рН раствора поддерживали в пределах 7.3-7.4, а температуру на уровне 200С. Препараты пищевода фиксировали вертикально одним концом к тензометрическому датчику, другим - к неподвижному штативу, а затем погружали в отдельные резервуары объемом 20 мл, куда подавался перфузионный раствор и карбоген. Сокращения полосок пищевода вызывали электрической стимуляцией с частотой 1 имп/мин, амплитудой сигнала 10 В, продолжительностью стимула 50 мс. После погружения препаратов в резервуары следовал период приработки в течение 40-60 минут, в ходе которого мышечным полоскам постепенно придавалось оптимальное натяжение. Для исследования роли P2Y-рецепторов использовали следующие препараты: РС2 - антагонист P2Y2,4,6,11,13-рецепторов, PPADS - антагонист P2Y1,4,13-рецепторов, АТФ - агонист P2Y1,2,4,11,12,13-рецепторов, УТФ – агонист P2Y2,4,6,11 –рецепторов. Все использованные вещества – производства Sigma-Aldrich (США). Эффекты препаратов на сократительную активность оценивался через 60 минут от момента подачи в экспериментальную ванночку, концентрация – 10-4 М.
Обработка и статистический анализ данных
Толщину волокон циркулярного и продольного мышечных слоев пищевода крыс контрольной и ахалазийной групп оценивали на парафиновых срезах, окрашенных гематоксилин-эозином. Толщину вычисляли при помощи программы ImagePro как расстояние между противолежащими краями мышечного волокна. Регистрация сократительных ответов полосок пищевода и обработка полученных результатов проводилась с помощью программы Chart 5.55 (AD Instruments, США). Результаты исследования подвергались статистической обработке на персональном компьютере с использованием программных комплексов Origin 7.5. Все результаты представлены в виде М±m, где М – среднее значение, m – стандартная ошибка. Достоверность различий между средними величинами оценивалась с использованием t-критерия Стьюдента.

Результаты исследования и их обсуждение
Сравнительное изучение экспрессии P2Y1,2,4-рецепторов в пищеводе человека в норме и при ахалазии
Иммуногистохимические исследования P2Y1-рецепторов в контрольной группе показали, что позитивная реакция локализуется во всех слоях клеток многослойного плоского неороговевающего эпителия. Позитивная иммуногистохимическая реакция определяется в клетках, предположительно, в фибробластах подслизистого и шванновских клетках межмышечного сплетений, а также в нервных волокнах в толще мышечной стенки. Локализация P2Y2-рецепторов наблюдается в единичных клетках (фибробластах) подслизистого и шванновских клетках межмышечного слоев, в многослойном плоском эпителии, а также в волокнах внутреннего мышечного слоя стенки пищевода. P2Y4-рецепторы локализуются в многослойном плоском неороговевающем эпителии, в клетках подслизистого сплетения, во внутреннем и наружном мышечных слоях стенки пищевода. По данным зарубежных авторов в подслизистом сплетении органов ЖКТ присутствуют фибробласты, в которых экспрессируются P2Y1,2-рецепторы (Furuya K., 2005). Учитывая эти данные, следует предположить, что в подслизистом слое пищевода человека исследованные нами пуринорецепторы локализуются именно в фибробластах. Мы предполагаем, что зафиксированное окрашивание в межмышечном сплетении пищевода локализуется в шванновских клетках на основании сравнительного анализа иммуногистохимических результатов по экспрессии белка S100 и P2Y-рецепторов в стенке пищевода контрольной группы: S100, как и P2Y-рецепторы экспрессируются в нервных волокнах, а также в клетках округлой и колечкообразной формы преимущественно межмышечного сплетения, по структуре идентичных шванновским. Также, по данным мировой литературы P2Y-рецепторы преимущественно локализуются в нейронах, шванновских клетках и клетках Кахаля межмышечного сплетения органов ЖКТ (Giaroni C. et al., 2002, Van Nassauw L., 2006, Gallego D., 2008).
Нами выявлена зависимость между изменением экспрессии P2Y-рецепторов и видом лечения, которое получали больные с ахалазией. Так, у больных, не получавших какого-либо лечения до оперативного вмешательства, а также у пациентов, прошедших процедуру бужирования (пневмокардиодилатацию), на различных участках биоптата наблюдается яркая позитивная реакция к P2Y1-рецепторам во множественных толстых мышечных пучках и единичных тонких волокнистых структурах, которые могут являться дегенерировавшими нервными волокнами (остаточными бюнгнеровскими лентами). У больных, получавших до операции лечение ботулотоксином типа А (БТА), также наблюдается позитивная реакция в бюнгнеровских лентах, но в меньшей степени. Этот факт уменьшения экспрессии P2Y1-рецепторов можно объяснить тем, что мышечные волокна парализованы БТА и изначально находятся в расслабленном состоянии, поэтому адаптационный механизм к хроническому спазму выражен слабее.
Исследования P2Y2-рецепторов в пищеводе больных с ахалазией, прошедших процедуру бужирования и не получавших лечения до операции, показали, что в обоих случаях позитивная иммуногистохимическая реакция локализуется в волокнистых структурах (предположительно, в фиброзно-соединительных прослойках) рядом с ГМК и в пучках мышечных волокон. Следует предположить, что ГМК пищевода этих пациентов способны сокращаться. Усиление экспрессии P2Y1,2-рецепторов в пищеводе может являться адаптационным механизмом ткани в ответ на длительный спазм и механическую травму.
Исследования P2Y4-рецепторов в пищеводе больных с ахалазией показали, что наблюдается негативная реакция в пищеводах больных, получавших и не получавших лечение до операции. Таким образом, экспрессия P2Y1,2,4 –рецепторов широко представлена в стенке пищевода человека контрольной группы.
Морфофункциональный анализ модели экспериментальной ахалазии у крыс
ЭА пищевода у крыс с клинической и морфологической точки зрения характеризуется: 1) снижением массы тела крыс с ЭА к 20 суткам до 71.6±1.8 % от значений, полученных до операции (n=9, р<0.05); 2) повышенным давлением (гипертонусом) в области НПС и нижней трети пищевода на 14% (n=5, р<0.05) и 12% (n=5, р<0.05) при сравнении с контролем, соответственно; 3) пролиферацией и гиперплазией клеток многослойного плоского неороговевающего эпителия; 4) значительной гипертрофией (в 4 раза) продольных и циркулярных мышечных волокон НПС и нижней трети пищевода; 5) исчезновением экспрессии белка S100 в подслизистом нервном сплетении и регенерацией тонких волокнистых структур с формированием бюнгнеровских лент в межмышечном нервном сплетении; 6) разрастанием фиброзно-соединительной ткани в стенке пищевода. Таким образом, морфологическая картина пищевода при ЭА у крысы коррелирует с таковой при АП у человека, что подтверждает адекватность исследованной нами модели заболевания.
Сократительная активность пищевода крысы в норме и при экспериментальной ахалазии
В экспериментах на интактных крысах было выявлено два типа сократительных ответов пищевода на электрический стимул – быстрый и следующий за ним медленный ответ (рис. 1Б). Амплитуда быстрого ответа, как для НПС, так и нижней трети пищевода, была достоверно больше в сравнении с медленным (p<0.05). В случае НПС медленные сократительные ответы наблюдались в 92.3% случаев, а быстрые – лишь в 38.5% экспериментов (n=13). Таким образом, для НПС более характерны медленные ответы (рис. 1А), поэтому в дальнейшем только они были подвергнуты анализу. В 7.7% случаев наблюдались спонтанные сократительные ответы (рис. 1В), по форме сходные с медленными, электрически вызванными сокращениями. Для нижней трети пищевода были характерны двухфазные сократительные ответы, состоящие из быстрой (присутствовала в 100% случаев) и медленной (присутствовала в 92.3% случаев) фаз. Спонтанная активность нижней трети пищевода была выявлена значительно чаще в сравнении с НПС – она наблюдалась в 46.2% случаев. Двухфазность электрически вызванной сократительной активности пищевода может быть объяснена сокращением ГМК под действием электрического стимула (прямое раздражение) и в ответ на выделение медиатора из нервных окончаний (непрямое раздражение).
На модели ЭА характер сократительной активности пищевода значительно изменялся: быстрые сократительные ответы НПС наблюдались чаще (46.2% экспериментов), а медленные – реже (69.2%, n=13), однако амплитуда сокращений достоверно не отличалась в сравнении с интактными животными (p<0.05). В 15.4% случаев наблюдались спонтанные медленные сократительные ответы, что больше в сравнении с данными, полученными на интактных крысах. Для нижней трети пищевода были характерны, как и в норме, двухфазные сократительные ответы. Однако частота встречаемости как быстрых, так и медленных ответов, была снижена в сравнении с нормой – до 92.3% и 76.9% соответственно.

Рис. 1. Сократительная активность пищевода интактной крысы
На графиках А и Б представлена механомиографическая запись электрически вызванных сократительных ответов НПС и нижней трети пищевода интактных крыс, соответственно (усреднения по 20 реализациям). Стрелками указаны моменты нанесения электрических стимулов (ЭС). На графике В показан пример спонтанной сократительной активности нижней трети пищевода интактных крыс: общая длительность окна – 9 минут; частота активности - 3.2 мин-1. Данные отдельных экспериментов.
Амплитуда быстрого ответа была достоверно выше в сравнении с данными, полученными на интактных крысах (p<0.05), тогда как амплитуда медленного ответа не отличалась (p>0.05). Спонтанная активность нижней трети пищевода при ахалазии была меньше по амплитуде и встречалась значительно реже в сравнении с контрольными показателями – лишь в 30.8% экспериментов.
Таким образом, мы впервые выявили характер нарушений сократительной активности пищевода на модели АП. При ЭА в случае НПС в сравнении с нормой чаще встречались быстрые вызванные ответы и спонтанная активность, что может объясняться растормаживанием ГМК вследствие дегенерации тормозных нейронов пищевода. Снижение частоты встречаемости медленных вызванных ответов может, по всей видимости, быть связано с нейромедиаторным дефицитом в результате дегенерации нейронов (Takahashi T., 2003), а также с повышенным тонусом НПС при АП.
Сравнительное изучение экспрессии P2Y1,2,4-рецепторов в стенке пищевода крыс контрольной группы и при экспериментальной ахалазии
Иммуногистохимические исследования P2Y1-рецепторов в контрольной группе крыс показали, что яркая позитивная реакция наблюдается в многослойном плоском неороговевающем эпителии, а также в волокнах циркулярного мышечного слоя. P2Y2-рецепторы локализуются в многослойном плоском неороговевающем эпителии, нервных волокнах межмышечного нервного сплетения, продольном мышечном слое стенки пищевода. P2Y4-рецепторы локализуются в многослойном плоском неороговевающем эпителии пищевода, клетках, предположительно, фибробластах подслизистого и шванновских клетках межмышечного нервных сплетений пищевода, в циркулярных и продольных мышечных волокнах. Иммуногистохимические исследования P2Y1-рецепторов у крыс с ЭА показали, что сохраняется позитивная реакция в мышечных волокнах внутреннего (циркулярного слоя) и в многослойном плоском эпителии. Исследования P2Y2-рецепторов у крыс с ЭА выявили, что позитивная иммуногистохимическая реакция локализуется, в отличие от контрольной группы, в циркулярных мышечных волокнах и исчезает в многослойном плоском неороговевающем эпителии. Изучение P2Y4-рецепторов у крыс опытной группы показало, что экспрессия данного типа пуринорецепторов отсутствует во всех слоях многослойного плоского неороговевающего эпителия и не наблюдается в гладкомышечных волокнах циркулярного и продольного слоев. Сопоставляя результаты, полученные по локализации P2Y1,2,4-рецепторов в пищеводе человека и крыс в контроле и при ахалазии наблюдается схожая картина по локализации указанных типов P2Y-рецепторов.
Роль P2Y-рецепторов в регуляции сократительной активности пищевода в норме и при экспериментальной ахалазии
В целях изучения участия P2Y1,2,4-рецепторов в регуляции сократительной активности пищевода использовали ряд наиболее распространенных антагонистов и агонистов P2Y-рецепторов. В норме, в случае НПС, неспецифический блокатор P2Y-рецепторов РС2 приводил к увеличению амплитуды медленного ответа до 325.3±35.8% от исходной величины (n=4, p<0.05). Более специфический блокатор PPADS оказывал сходное, но гораздо менее выраженное влияние - увеличивал амплитуду сокращений до 125.1±10.3% от исходной (n=4), (рис. 2).
Активаторы пуринорецепторов АТФ и УТФ не оказывали достоверного влияния на амплитуду медленных сократительных ответов (p>0.05).
Полученные нами данные об эффектах P2Y-антагонистов свидетельствуют о наличии P2Y-опосредованной тонической активности тормозных нейронов, ингибирующих в норме сократительную активность НПС. Сопоставляя эффекты различных агонистов и антагонистов пуринорецепторов, можно говорить о наибольшем участии P2Y2- P2Y4- и P2Y6-рецепторов в регуляции тонуса НПС, а также об отсутствии существенного вклада со стороны P2X-рецепторов.
При ЭА в случае НПС, неспецифический антагонист P2Y-рецепторов РС2 достоверно не влиял на амплитуду медленного ответа (n=4, p>0.05). Более специфический блокатор P2Y-рецепторов PPADS вызывал снижение амплитуды медленных ответов до 75.7±9.5 % от начального значения (n=4, p<0.05), (рис. 2). Таким образом, при ЭА эффекты P2Y-антагонистов значительно видоизменялись – отсутствовали (РС2), либо имели противоположную направленность (PPADS).
Эффекты агонистов пуринорецепторов также значительно отличались. Как АТФ, так и УТФ в условиях ЭА приводили к усилению амплитуды медленных ответов НПС – до 139.4±18.4 и 170.3±28.1% от исходных значений соответственно (n=4, p<0.05).
Значительная модификация эффектов P2Y-агонистов и антагонистов при ЭА может объясняться отсутствием P2Y-опосредованного нейронального контроля в результате гибели тормозных нейронов. В этом случае будут преобладать эффекты на P2Y-рецепторы непосредственно ГМК. Более выраженные влияния PPADS в сравнении с РС2 и УТФ в сравнении с АТФ свидетельствуют об увеличении экспрессии P2Y1-рецепторов, в отношении которых РС2 и АТФ неактивны. Это наблюдение подтверждается данными иммуногистохимии (неопубликованные данные). Примечателен тот факт, что АТФ и УТФ усиливают сократительную активность НПС при ЭА, но не в норме. Возможно, этот феномен оказывает вклад в патогенетический механизм развития АП.
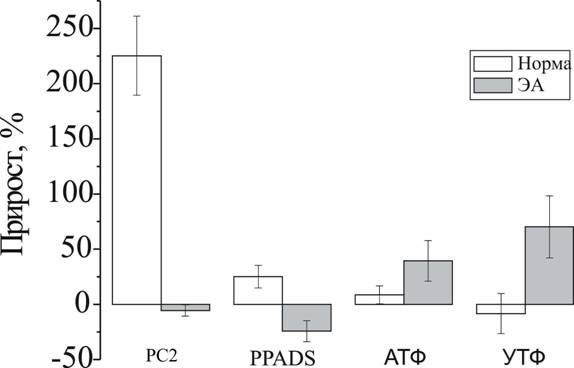
Рис. 2. Влияние P2Y-агонистов и антагонистов на сократительную активность нижнего пищеводного сфинктера.
Показаны изменения амплитуды медленных сократительных ответов НПС под действием P2Y-антагонистов (РС2, PPADS) и агонистов (АТФ, УТФ) через 60 минут от начала действия вещества в норме (белые столбцы) и при экспериментальной ахалазии (ЭА, серые столбцы)
Выводы
1. В нижней трети пищевода человека контрольной группы P2Y1,2,4-рецепторы локализуются в клетках многослойного плоского неороговевающего эпителия, в фибробластах подслизистого слоя, шванновских клетках и нервных волокнах межмышечного нервного сплетения, в продольных и циркулярных мышечных волокнах.
2. В биоптатах пищевода пациентов с ахалазией меняется локализация P2Y1,2,4-рецепторов: происходит исчезновение P2Y4-рецепторов в мышечной стенке, сохраняется экспрессия P2Y1-рецепторов в мышечных волокнах и тонких волокнистых структурах (бюнгнеровских лентах), P2Y2-рецепторов – в фиброзно-соединительных прослойках между гладкомышечными волокнами.
3. В нижней трети стенки пищевода крысы в норме P2Y1,2,4-рецепторы экспрессируются в клетках многослойного плоского неороговевающего эпителия, в фибробластах подслизистого нервного сплетения, в шванновских клетках и нервных волокнах межмышечного нервного сплетения, в продольных и циркулярных мышечных волокнах.
4. В нижней трети стенки пищевода крыс с экспериментальной ахалазией меняется локализация P2Y-рецепторов: P2Y2–рецепторы локализуются в волокнах циркулярного мышечного слоя и исчезают в многослойном плоском эпителии и в продольном мышечном слое, полностью отсутствует экспрессия P2Y4-рецепторов во всех гистологических структурах.
5. Морфологическая картина нижней трети пищевода крыс с экспериментальной ахалазией соответствует таковой при ахалазии у человека и характеризуется: 1) пролиферацией клеток многослойного плоского неороговевающего эпителия с гиперплазией до 15 слоев; 2) гипертрофией циркулярных и продольных мышечных волокон нижней трети пищевода; 3) отсутствием экспрессии белка S100 в подслизистом нервном сплетении и ее наличием в бюнгнеровских лентах в межмышечном нервном сплетении; 4) значительным увеличением экспрессии виментина между мышечными волокнами и в подслизистом слое по сравнению с контрольной группой.
6. В миографических исследованиях сократительной активности нижнего пищеводного сфинктера в норме выявлено два типа сократительных ответов – быстрый и следующий за ним медленный ответ (время нарастания 4.77±1.03 с, время полурасслабления - 4.81±0.88 с). Медленные сократительные ответы наблюдались в 2.4 раза чаще в сравнении с быстрыми. При экспериментальной ахалазии частота встречаемости быстрых вызванных ответов увеличивалась (46.2% экспериментов), а медленных – уменьшалась (69.2%).
7. В миографических исследованиях сократительной активности нижней трети пищевода в контроле выявлены двухфазные сократительные ответы, состоящие из быстрой и медленной фаз. В отличие от нижнего пищеводного сфинктера, быстрая фаза присутствовала в 100% случаев (время нарастания - 0.27±0.02 с., время расслабления - 0.3±0.03 с.), а медленная фаза - в 92.3% случаев (время нарастания - 3.42±0.43 с., время полурасслабления - 4.94±0.9 с.). Спонтанная активность нижней трети пищевода наблюдалась в 7 раз чаще в сравнении с нижним пищеводным сфинктером. При экспериментальной ахалазии частота встречаемости как быстрых (в 92.3% экспериментов) так и медленных (в 76.9% экспериментов) ответов была снижена в сравнении с контролем. Спонтанная активность была меньше по амплитуде и встречалась значительно реже в сравнении с контрольными показателями.
8. В контрольных условиях блокирование метаботропных P2Y-рецепторов под действием PPADS или РС2 приводило к увеличению амплитуды медленных сократительных ответов нижнего пищеводного сфинктера крысы – до 325.3±35.8% и 125.1±10.3% от исходных значений, соответственно. Эффекты активаторов P2Y-рецепторов АТФ и УТФ не оказывали достоверного влияния на амплитуду медленных сократительных ответов.
9. В условиях экспериментальной ахалазии эффекты агонистов и антагонистов P2Y-рецепторов на амплитуду медленных ответов нижнего пищеводного сфинктера значительно видоизменялись в сравнении с контролем: РPADS вызывал ее снижение до 75.7±9.5 % от исходного значения, РС2 не оказывал достоверного влияния, АТФ и УТФ приводили к увеличению амплитуды – до 139.4±18.4 и 170.3±28.1% от исходных величин, соответственно.
10. Эффекты как агонистов, так и антагонистов P2Y-рецепторов не оказывали достоверного влияния на быстрые и медленные сократительные ответы нижней трети пищевода.
Практические рекомендации
Рекомендовать внедрение в практику патологоанатомических отделений диагностические мероприятия в виде иммуногистохимической оценки экспрессии P2Y1,2,4-рецепторов в биоптатах стенки пищевода больных ахалазией с целью дальнейшего подбора и коррекции лечения пациентов с данным заболеванием при помощи лекарственных препаратов с пуринергическим механизмом действия.
Список работ, опубликованных по теме диссертации
1. Вклад P2Y-рецепторов в регуляцию сократительной активности пищевода in vivo и in vitro / [и др.] // Биология – наука XXI века: тез. докл. 12-ой междунар. школы-конференции молодых ученых. – Пущино, 2008. – С. 149-150.
2. Пуринэргический контроль сократительной активности пищевода в норме и в модели ахалазии у крыс / [и др.] // Структурные и функциональные основы эволюции функций, физиология экстремальных состояний: тез. докл. Всерос. конф. «Научное наследие академика ». – СПб., 2008. – С. 106-107.
3. Влияние P2Y-рецепторов на моторику пищевода / [и др.] // Структурные и функциональные основы эволюции функций, физиология экстремальных состояний: тез. докл. Всерос. конф. «Научное наследие академика ». – СПб., 2008. – С. 143-144.
4. Применение препарата «Ксимедон» в лечении экспериментальной ахалазии у крыс / [и др.] // Фармакология и токсикология фосфорорганических соединений и других биологически активных веществ: тез. докл. Рос. конф., посвящ. 75-летию проф. . – Казань, 2008. – С. 46-47.
5. Модификация метода экспериментальной ахалазии у крыс и анализ его эффективности / [и др.] // Казанский медицинский журнал. – 2009. – Т. 90, №2. – С. 282-284.
6. Развитие экспериментальной ахалазии пищевода под влиянием производного пиримидина ксимедона / [и др.] // Актуальные проблемы патофизиологии: тез. докл. XV межгородской конф. молодых ученых. – СПб., 2009. – С. 95-96.
7. Сабиров регуляция сократительной активности пищевода в норме и в модели ахалазии / // Молодые ученые в медицине: тез. докл. XIV Всерос. науч.-практ. конф. с междунар. участием, Казань, 29-30 апр. 2009 г. / науч. ред.: . – Казань, 2009. – С. 174.
8. Морфофункциональный анализ экспериментальной модели ахалазии пищевода у крыс / [и др.] // Бюллетень экспериментальной биологии и медицины. – 2010. – Т. 148, №4. – С. 452-456.
9. Пуринергическая регуляция сократительной активности пищевода в норме и в условиях экспериментальной ахалазии / [и др.] // Молодые ученые в медицине: тез. докл. XIV Всерос. науч.-практ. конф. с междунар. участием. – Казань, 2010. – С. 287.
10. Диагностика и лечение ахалазии кардии: учеб. пособие для врачей / [ и др.]. – Казань: [б. и.]. – 44 с.
Список сокращений
АП – ахалазия пищевода
АТФ - аденозин 5’-трифосфорная кислота
БТА - ботулотоксин типа А
БХ - бензалкония хлорид
ГМК – гладкомышечные клетки
НПС – нижний пищеводный сфинктер
РКОД МЗ РТ - Республиканский Клинический Онкологический Диспансер Министерства Здравоохранения Республики Татарстан
PC2 – реактив синий 2
УТФ - уридин-5′трифосфорная кислота
ЭА – экспериментальная ахалазия
P2Y – метаботропные пуринорецепторы
P2X - ионотропные рецепторы
PPADS - пиридоксальфосфат-6-азафенил-2’,4’-дисульфоновая кислота
