

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
В процесса индивидуального развития организм закономерно меняет свои характеристики. Особенно интенсивные изменения происходят в эмбриональном периоде онтогенеза, когда из зиготы формируются структуры многоклеточного организма. При этом все многообразие клеток, выполняющих в организме различные функции, происходит из одной клетки путем митотического деления. Так как в результате митоза дочерние клетки получают полноценную наследственную информацию, заключенную в кариотипе, все клетки организма в генотипическом отношении равноценны. Некоторые различия, однако, наблюдаются за счет цитоплазматических генов, например митохондриальных, которые распределяются при делении не строго равномерно.
Чем определяются морфологические, физиологические и биохимические различия, появляющиеся между клетками в ходе развития? В процессе овогенеза в цитоплазме яйцеклетки накапливаются не только богатые энергией вещества, обеспечивающие развитие зародыша, но и мРНК для синтеза белков, необходимых на самых ранних стадиях эмбрионального развития. Распределение этих веществ в цитоплазме яйцеклетки оказывается неравномерным. Проникновение сперматозоида в яйцеклетку вызывает перераспределение отдельных компонентов в объеме клетки, в связи с чем уже при первых делениях зиготы в дочерних клетках оказывается цитоплазма с разным составом веществ.
Взаимодействие между компонентами цитоплазмы и ядром приводит к дерепрессии определенных генов. Их продукты определяют дальнейшее углубление различий между разными частями зародыша, т. е. дифференцировку. Возникающие различия порождают новые взаимодействия между соседними клеточными группами, которые вызывают дерепрессию новых генов, вследствие чего меняется спектр активных генов и, следовательно, генетическая программа на последующий отрезок процесса развития. Таким образом, в ходе индивидуального развития первоначально репрессированный геном зиготы подвергается постепенной дерепрессии, причем в разных частях зародыша дерепрессируются разные группы генов. Набор активно функционирующих генов определяет своеобразие спектра белков, которые синтезируются клетками, выполняющими различные функции.
Как отмечалось выше, в процессе онтогенеза в клетках организма происходит смена активно функционирующих генов. Гены, транскрибировавшиеся в эмбриональном периоде, к моменту рождения или непосредственно после него репрессируются, в то же время активируются гены, определяющие специфические функции клеток во взрослом организме. Нередко вещества, продуцируемые определенным типом клеток в разные периоды онтогенеза, несколько различаются по своим свойствам. Изменение свойств диктуется изменением условий существования организма, например в эмбриональном и постэмбриональном периодах развития. Эти различия объясняются сменой функционирования близких, но не идентичных по заключенной в них информации генов. Такие гены в ряде случаев образуют группы, получившие название мультигенных семейств. Примером тому служат гены гемоглобина.
Мультигенное семейство — это группа генов, очень близких по нуклеотидным последовательностям, со сходными фенотипическими функциями. Число генов в разных семействах у представителей разных видов варьирует от единиц до нескольких сотен. К примеру, число генов гистонов у разных видов в отдельных семействах колеблется от 01.01.01, генов тРНК — от 6 до 400, генов 5SPHK — от 200 до 24000, генов α-глобинов — от 1 до 5, (β-глобинов — от 2 до 7. К числу белков, кодируемых мультигенными семействами, кроме указанных выше, относятся актины и тубулины, играющие важную роль в подвижности клеток, коллагены соединительной ткани, некоторые белки клеточных мембран и сыворотки крови.
Мультигенные семейства могут быть организованы в геноме по-разному (рис. 6.4). Так, члены семейства идентичных генов РНК у человека располагаются в виде тандемных повторов, в которых структурные гены разделены некодирующими — спейсерными — участками. Однако гены семейства не всегда идентичны. Например, в семействах глобиновых генов тандемно сцеплены близкие, но не одинаковые гены. В случае гистоновых генов у некоторых видов тандемно повторяются целые пакеты (кластеры) неидентичных генов, определяющих синтез разных видов гистонов, что может быть описано формулой Н2А — НЗ —Н2В — Н4 — H1 —. Возможно также диффузное распределение генов семейства по нескольким хромосомам, что показано для генов актинов и тубулинов.
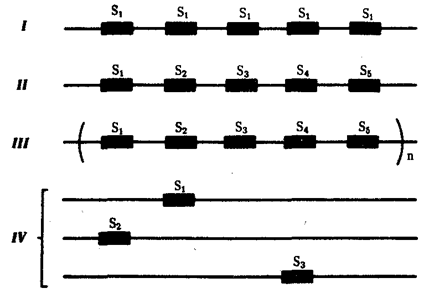
Рис. 6.4. Типы организации мультигенных семейств:
I — идентичные гены, сцепленные тандемно (гены рРНК); II — близкие, но не идентичные гены, сцепленные тандемно (гены глобинов); III — скопления неидентичных генов, сцепленных тандемно (гистоновые гены); IV — близкие гены, рассеянные по нескольким хромосомам (гены актинов или тубулинов)
Описанные варианты организации мультигенных семейств создают необходимые условия для эффективной регуляции экспрессии соответствующих генов. Так, если продукт определенного гена необходим лишь на небольшом отрезке времени в онтогенезе, но в значительных количествах, мультигенное семейство образовано большим числом идентичных генных копий, обычно соединенных тандемно. Примером могут служить гены рРНК, которые в геноме соматических клеток взрослой шпорцевой лягушки представлены 450 копиями. Вместе с тем в овогенезе для быстрого образования необходимого количества рибосом, которых в яйце Xenopus содержится около 1012, гены рРНК амплифицируются и число их копий возрастает в 4000 раз. В других мультигенных семействах, состоящих, как правило, из неидентичных генов, в ходе онтогенеза происходит переключение с одного гена на другой. Белки, контролируемые определенными генами такого семейства, наилучшим образом соответствуют либо условиям на разных стадиях онтогенеза, либо клеткам различных типов.
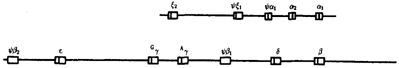
Рис. 6.5. Распределение генов α- и β-глобинов человека
Глобиновые гены изображены в виде прямоугольников; вертикальные полосы внутри соответствуют экзонам, остальные участки — интронам
Наиболее изучены в этом отношении мультигенные семейства α- и β-глобиновых генов (рис. 6.5). У человека они представлены кластером из 7 β-глобиновых генов, расположенных в 11-й хромосоме, и кластером из 5 α-глобиновых генов, локализующихся в 16-й хромосоме. У эмбрионов человека активно функционируют ζ (дзета)-глобиновый ген из α- семейства и ε (эпсилон)-глобиновый ген из β-семейства, обеспечивающие образование эмбрионального ζ2ε2-гемоглобина.
На более поздних стадиях онтогенеза у плода эти гены репрессируются, но дерепрессируются другие гены семейств, определяющие синтез фетального α2γ2-гемоглобина. После рождения начинают экспрессироваться δ (дельта)- β и (бета)-глобиновые гены, обеспечивающие образование преобладающего α2β2- и минорного α2β2- видов гемоглобина взрослого человека. В обоих кластерах имеются также неэкспрессирующиеся псевдогены ψα1, ψξ1, ψβ1, ψβ2.
Переключение генов в мультигенных семействах происходит не только в соответствии со стадией индивидуального развития, но и с типом и местом локализации клеток в организме. Так, эмбриональный гемоглобин С, г б2 образуется имеющими ядра мегалобластами в стенке желточного мешка. На 6-й неделе развития происходит смена экспрес-сируемых генов и переход к синтезу гемоглобина безъядерными эритроцитами печени и селезенки плода. Позднее главным местом образования гемоглобина становится костный мозг, где вскоре после рождения начинается синтез взрослых гемоглобинов α2β2 и α2δ2. Смена типов синтезируемого гемоглобина у эмбриона, плода и после рождения связана с конкретными условиями существования организма на разных стадиях онтогенеза. Так, у человека гемоглобин плода имеет более высокое сродство к кислороду, чем гемоглобин взрослого, что облегчает перенос кислорода через плаценту.
Таким образом, изменение характеристик фенотипа организма на разных стадиях онтогенеза является результатом регуляции экспрессии генов, которая имеет целью в одних случаях наращивание продукции определенных белков, а в других — переход от синтеза одного белка к синтезу белка более соответствующего изменяющимся условиям существования.
Наследственная программа, на основе которой формируется фенотип организма, сосредоточена главным образом в его хромосомном наборе. Некоторое количество наследственного материала заключено также в цитоплазме клеток. Ядерные и цитоплазматические структуры в процессе клеточного размножения распределяются между дочерними клетками по-разному. Это касается не только соматических клеток организма, но и его гамет. В связи с этим передача ядерных и цитоплазматических генов потомству подчиняется разным закономерностям, что обусловливает особенности наследования соответствующих признаков.
6.3.1. Закономерности наследования признаков,
контролируемых ядерными генами
Гены, расположенные в ядерных структурах — хромосомах, закономерно распределяются между дочерними клетками благодаря механизму митоза, который обеспечивает постоянную структуру кариотипа в ряду клеточных поколений (см. разд. 3.6.2.1). Мейоз и оплодотворение обеспечивают сохранение постоянного кариотипа в ряду поколений организмов, размножающихся половым путем (см. разд. 3.6.2.2). В результате набор генов, заключенный в кариотипе, также остается постоянным в ряду поколений клеток и организмов. Закономерное поведение хромосом в митозе, мейозе и при оплодотворении обусловливает закономерности наследования признаков, контролируемых ядерными генами.
6.3.1.1. Моногенное наследование признаков.
Аутосомное и сцепленное с полом наследование
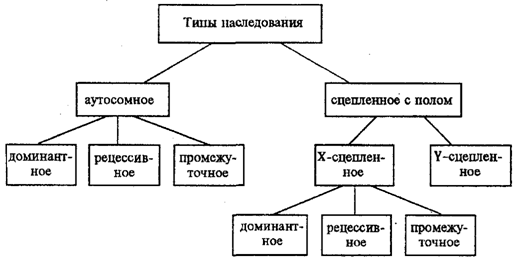
В связи с тем что кариотип организма — это диплоидный набор хромосом, большинство генов в соматических клетках представлены аллельными парами. Аллелъные гены, расположенные в соответствующих участках гомологичных хромосом, взаимодействуя между собой, определяют развитие того или иного варианта соответствующего признака (см. разд. 3.6.5.2). Являясь специфической характеристикой вида, кариотип представителей разного пола различается по паре половых хромосом (см. разд. 6.1.2.1).Гомогаметный пол, имеющий две одинаковые половые хромосомы XX, диплоиден по генам этих хромосом. Гетерогаметный пол имеет одинарный набор генов Х-хромосомы (ХО) или негомологичных участков Х - и Y-хромосом. Фенотипическое проявление и наследование отдельных признаков из поколения в поколение организмов зависит от того, в каких хромосомах располагаются соответствующие гены и в каких дозах они присутствуют в генотипах отдельных особей. Различают два основных типа наследования признаков: аутосомное и сцепленное с полом (схема 6.1).
Схема 6.1. Классификация типов наследования признаков
при моногенном наследовании
Аутосомное наследование. Характерные черты аутосомного наследования признаков обусловлены тем, что соответствующие гены, расположенные в аутосомах, представлены у всех особей вида в двойном наборе. Это означает, что любой организм получает такие гены от обоих родителей. В соответствии с законом чистоты гамет в ходе гаметогенеза все половые клетки получают по одному гену из каждой аллельной пары (рис. 6.6). Обоснованием этого закона является расхождение гомологичных хромосом, в которых располагаются аллельные гены, к разным полюсам клетки в анафазе I мейоза (см. рис. 5.6).
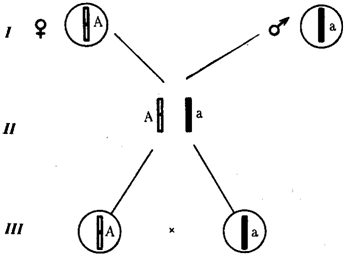
Рис. 6.6. Обоснование закономерностей аутосомного наследования признаков:
I — гаплоидные гаметы родителей, II — диплоидный генотип особи (фенотип зависит от взаимодействия аллельных генов Аа); III— гаплоидные гаметы гетерозиготной особи (гаметы «чисты», так как несут по одному из пары аллельных генов); черным и белым обозначены гомологичные хромосомы; буквами — определенные локусы
Ввиду того что развитие признака у особи зависит в первую очередь от взаимодействия аллельных генов, разные его варианты, определяемые разными аллелями соответствующего гена, могут наследоваться по аутосомно-доминантному или аутосомно-рецессивному типу, если имеет место доминирование. Возможен также промежуточный тип наследования признаков при других видах взаимодействия аллелей (см. разд. 3.6.5.2).
При доминировании признака, описанном Г. Менделем в его опытах на горохе, потомки от скрещивания двух гомозиготных родителей, различающихся по доминантному и рецессивному вариантам данного признака, одинаковы и похожи на одного из них (закон единообразия F1). Описанное Менделем расщепление по фенотипу в F2 в отношении 3:1 в действительности имеет место лишь при полном доминировании одного аллеля над другим, когда гетерозиготы фенотипически сходны с доминантными гомозиготами (закон расщепления в F2).
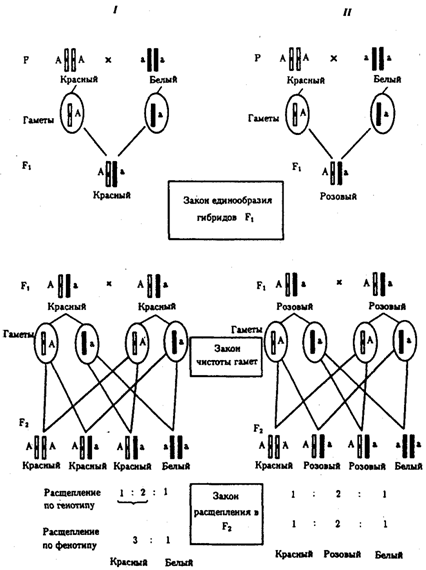
Рис. 6.7. Аутосомное наследование признака:
I — полное доминирование (наследование цвета лепестков у гороха); II — неполное доминирование (наследование цвета лепестков у ночной красавицы)
Наследование рецессивного варианта признака характеризуется тем, что он не проявляется у гибридов F1, а в F2 проявляется у четверти потомков (рис. 6.7.7).
В случаях формирования у гетерозигот нового варианта признака по сравнению с гомозиготами, что наблюдается при таких видах взаимодействия аллельных генов, как неполное доминирование, кодоминирование, межаллельная комплементация, гибриды F1 не похожи на родителей, а в F2 образуется три фенотипических группы потомков (рис. 6.7, II).
Завершая описание характерных черт аутосомно-доминантного и аутосомно-рецессивного наследования, уместно напомнить, что хотя в случае доминирования одного из аллелей присутствие в генотипе другого, рецессивного, аллеля не сказывается на формировании доминантного варианта признака, на фенотипическое проявление аллеля оказывает влияние вся система генотипа конкретного организма, а также среда, в которой реализуется наследственная информация. В связи с этим существует возможность неполной пенетрантности доминантного аллеля у особей, имеющих его в генотипе.
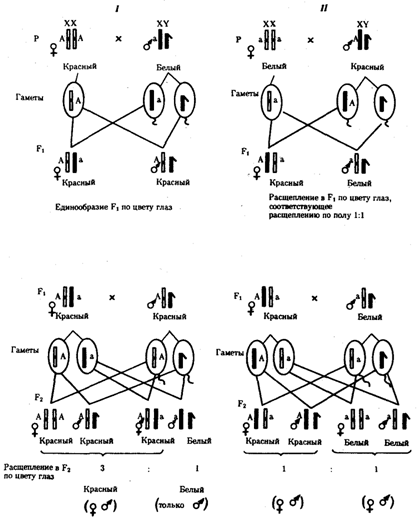
Рис. 6.8. Наследование признака окраски глаз у дрозофилы:
I, II — различие результатов скрещивания в зависимости от пола родителя с доминантным признаком; зачернены отцовские хромосомы
Сцепленное с полом наследование. Анализ наследования признака окраски глаз у дрозофилы в лаборатории Т. Моргана выявил некоторые особенности, заставившие выделить в качестве отдельного типа наследования признаков сцепленное с полом наследование (рис. 6.8).
Зависимость результатов эксперимента от того, кто из родителей являлся носителем доминантного варианта признака, позволила высказать предположение, что ген, определяющий окраску глаз у дрозофилы, расположен в Х-хромосоме и не имеет гомолога в Y-хромосоме. Все особенности сцепленного с полом наследования объясняются неодинаковой дозой соответствующих генов у представителей разного — гомо - и гетерогаметного пола.
Гомогаметный пол несет двойную дозу генов, расположенных в Х-хромосоме. Развитие соответствующих признаков у гетерозигот (ХAХa) зависит от характера взаимодействия между аллельными генами. Гетерогаметный пол имеет одну Х-хромосому (ХО или XY). У некоторых видов Y-хромосома генетически инертна, у других она содержит некоторое количество структурных генов, часть из которых гомологична генам Х-хромосомы (рис. 6.9). Гены негомологичных участков Х - и Y-хромосом (или единственной Х-хромосомы) у гетерогаметного пола находятся в гемизиготном состоянии. Они представлены единственной дозой: ХAY, ХaХ, XYB. Формирование таких признаков у гетерогаметного пола определяется тем, какой аллель данного гена присутствует в генотипе организма.
Характер наследования сцепленных с полом признаков в ряду поколений зависит от того, в какой хромосоме находится соответствующий ген. В связи с этим различают Х-сцепленное и Y-сцепленное (голандрическое) наследование.
Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х-хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству.
У млекопитающих (в том числе и человека) мужской пол получает Х-сцепленные гены от матери и передает их дочерям. При этом мужской пол никогда не наследует отцовского Х-сцепленного признака и не передает его своим сыновьям (рис. 6.10).
Так как у гомогаметного пола признак развивается в результате взаимодействия аллельных генов, различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. Х-сцепленный доминантный признак (красный цвет глаз у дрозофилы) передается самкой всему потомству. Самец передает свой Х-сцепленный доминантный признак лишь самкам следующего поколения. Самки могут наследовать такой признак от обоих родителей, а самцы — только от матери.
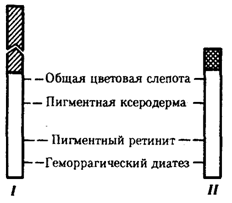
Рис. 6.9. Схема гомологичных и негомологичных локусов
в половых хромосомах человека:
I — Х-хромосомы: заштрихованы локусы, отсутствующие в Y-хромосоме (красно-зеленая слепота, гемофилия и др.); II — Y-хромосома: заштрихованы локусы, отсутствующие в Х-хромосоме (перепонки между пальцами, гены-детерминаторы развития организма по мужскому типу); участки Х - и Y-хромосом, соответствующие гомологичным локусам, не заштрихованы
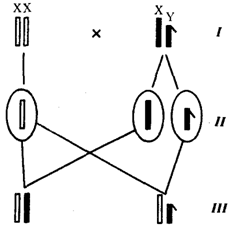
Рис. 6.10. Обоснование закономерностей сцепленного с полом
наследования признаков:
I — сочетание половых хромосом в кариотипе представителей разного пола; II — гомогаметный пол образует один тип гамет, гетерогаметный — два; III — представители гомогаметного пола получают хромосомы от обоих родителей; представители гетерогаметного пола получают Х-хромосому от гомогаметного родителя, а Y-хромосому — от гетерогаметного родителя; это справедливо для генов, расположенных в негомологичных локусах Х - и Y-хромосом; зачернены отцовские хромосомы
Х-сцепленный рецессивный признак, (белый цвет глаз у дрозофилы) у самок проявляется только при получении ими соответствующего аллеля от обоих родителей (XaXa). У самцов XaY он развивается при получении рецессивного аллеля от матери. Рецессивные самки передают рецессивный аллель потомкам любого пола, а рецессивные самцы —только «дочерям» (см. рис. 6.8).
При Х-сцепленном наследовании, так же как и при аутосомном, возможен промежуточный характер проявления признака у гетерозигот. Например, у кошек пигментация шерсти контролируется Х-сцепленным геном, разные аллели которого определяют черную (XA и рыжую (XA’) пигментацию. Гетерозиготные самки XAXA’ имеют пеструю окраску шерсти. Самцы же могут быть либо черными (XAY, либо рыжими (XA’Y).
Голандрическое наследование. Активно функционирующие гены Y-хромосомы, не имеющие аллелей в Х-хромосоме, присутствуют в генотипе только гетерогаметного пола, причем в гемизиготном состоянии. Поэтому они проявляются фенотипически и передаются из поколения в поколение лишь у представителей гетерогаметного пола. Так, у человека признак гипертрихоза ушной раковины («волосатые уши») наблюдается исключительно у мужчин и наследуется от отца к сыну.
6.3.1.2. Одновременное наследование нескольких признаков. Независимое и сцепленное наследование
Ранее были рассмотрены характерные черты фенотипического проявления и наследования отдельных признаков. Однако фенотип организма представляет собой совокупность многих свойств, за формирование которых отвечают разные гены. Так как общее число генов в генотипе значительно больше числа хромосом, каждая хромосома заключает в себе комплекс генов. В связи с этим неаллельные гены могут располагаться либо в разных хромосомах, либо входить в состав одной из них, занимая разные локусы АВС, КМР. Этим определяется характер наследования группы признаков, которое может быть независимым или сцепленным.
Независимое наследование признаков. Такой характер наследования признаков впервые был описан Г. Менделем в опытах на горохе, когда одновременно анализировалось наследование в ряду поколений нескольких признаков, например цвета и формы горошин (рис. 6.11). Каждый из них в отдельности подчинялся закону расщепления в F2. В то же время разные варианты этих признаков свободно комбинировались у потомков, встречаясь как в сочетаниях, наблюдаемых у их родителей (желтый цвет и гладкая форма или зеленый цвет и морщинистая форма), так и в новых сочетаниях (желтый цвет и морщинистая форма или зеленый цвет и гладкая форма). На основании анализа полученных результатов Г. Мендель сформулировал закон независимого наследования признаков, в соответствии с которым: «Разные пары признаков, определяемые неаллельными генами, передаются потомкам независимо друг от друга и комбинируются у них во всех возможных сочетаниях».
Очевидно, этому закону должны подчиняться в первую очередь неаллельные гены, располагающиеся в разных (негомологичных) хромосомах. В таком случае независимый характер наследования признаков объясняется закономерностями поведения негомологичных хромосом в мейозе. Названные хромосомы образуют со своими гомологами разные пары, или биваленты, которые в метафазе I мейоза случайно выстраиваются в плоскости экватора веретена деления. Затем в анафазе I мейоза гомологи каждой пары расходятся к разным полюсам веретена независимо от других пар. В результате у каждого из полюсов возникают случайные сочетания отцовских и материнских хромосом в гаплоидном наборе (см. рис. 3.75). Следовательно, различные гаметы содержат разные комбинации отцовских и материнских аллелей неал-лельных генов.
Разнообразие типов гамет, образуемых организмом, определяется степенью его гетерозиготности и выражается формулой 2n, где n — число локусов в гетерозиготном состоянии. В связи с этим дигетерозиготные гибриды F1 образуют четыре типа гамет с одинаковой вероятностью. Реализация всех возможных встреч этих гамет при оплодотворении приводит к появлению в F2 четырех фенотипических групп потомков в соотношении 9:3:3:1. Анализ потомков F2 по каждой паре альтернативных признаков в отдельности выявляет расщепление в соотношении 3:1.
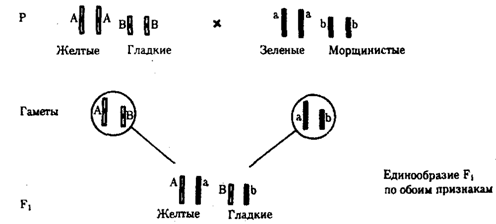
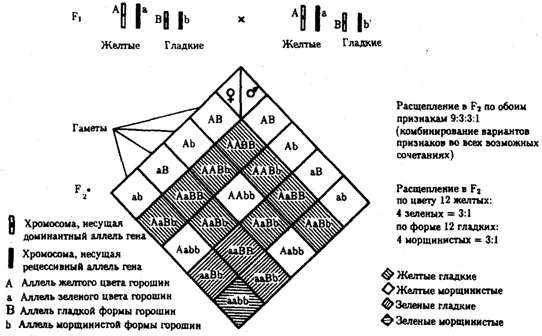
Рис. 6.11. Независимое наследование признаков (цвета и формы горошин)
Открытие независимого характера наследования разных признаков у гороха дало возможность Г. Менделю высказать предположение о дискретности наследственного материала, в котором за каждый признак отвечает своя пара наследственных задатков, сохраняющих в ряду поколений свою структуру и не смешивающихся друг с другом. Современные представления о надмолекулярной организации наследственного материала в хромосомах и закономерностях их передачи в ряду поколений клеток и организмов объясняют независимый характер наследования признаков расположением соответствующих генов в негомологичных хромосомах.

Рис. 6.12. Анализирующее (моногибридное) скрещивание
Объяснение см. в тексте
Анализирующее скрещивание. В опытах Г. Менделя наследственная конституция гибридов F1 устанавливалась на основе анализа результатов их самоопыления, получаемых в F2. В настоящее время с целью выяснения генотипа организмов с доминантным фенотипом (гомо- или гетерозигота) широко применяют также анализирующее скрещивание. Оно заключается в скрещивании организма, генотип которого необходимо определить, с организмом, несущим рецессивный признак, а следовательно, гомозиготным по рецессивному аллелю (рис. 6.12).
Так как гомозиготные организмы образуют один тип гамет: аа — (a), aabb — (ab), aabbcc — (abc) и т. д., — при анализирующем скрещивании количество фенотипов потомков зависит от числа типов гамет, образуемых организмом с доминантным фенотипом. Если последний гомозиготен по анализируемым генам, то он также образует только один тип гамет и потомство от анализирующего скрещивания единообразно и имеет доминантный фенотип (рис. 6.12, I).
Если анализируемый организм гетерозиготен по одному гену, он образует два типа гамет и при анализирующем скрещивании появляются потомки двух разных фенотипов с доминантным или рецессивным признаком (рис. 6.12, II).
Дигетерозиготный организм при анализирующем скрещивании дает четыре вида потомков (рис. 6.13).
В том случае, когда неаллельные гены А и В наследуются независимо, располагаясь в разных хромосомах, Дигетерозиготный организм образует четыре типа гамет с равной вероятностью. Поэтому в результате анализирующего скрещивания четыре фенотипически различающиеся вида потомков появляются в соотношении 1:1:1:1 и несут различные сочетания вариантов двух признаков.
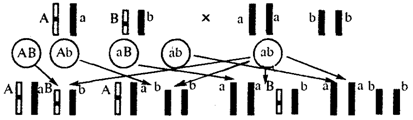
Рис. 6.13. Анализирующее (дигибридное) скрещивание Объяснение см. в тексте
Сцепленное наследование признаков. Анализ наследования одновременно нескольких признаков у дрозофилы, проведенный Т. Морганом, показал, что результаты анализирующего скрещивания гибридов Fi иногда отличаются от ожидаемых в случае их независимого наследования. У потомков такого скрещивания вместо свободного комбинирования признаков разных пар наблюдали тенденцию к наследованию преимущественно родительских сочетаний признаков. Такое наследование признаков было названо сцепленным. Сцепленное наследование объясняется расположением соответствующих генов в одной и той же хромосоме. В составе последней они передаются из поколения в поколение клеток и организмов, сохраняя сочетание аллелей родителей.
Зависимость сцепленного наследования признаков от локализации генов в одной хромосоме дает основание рассматривать хромосомы как отдельные группы сцепления.
На рис. 6.14 представлены результаты анализа наследования признаков окраски тела и формы крыльев у дрозофилы, а также их цитологическое обоснование. Обращает внимание, что при анализирующем скрещивании самцов из F1 появлялось всего два вида потомков, сходных с родителями по сочетанию вариантов анализируемых признаков (серая окраска тела и нормальные крылья или черная окраска тела и короткие крылья) в соотношении 1:1. Это указывает на образование самцами F1 всего двух типов гамет с одинаковой вероятностью, в которые попадают исходные родительские сочетания аллелей генов, контролирующих названные признаки (BV или bv).
При анализирующем скрещивании самок F1 появлялось четыре вида потомков со всеми возможными сочетаниями признаков. При этом потомки с родительскими сочетаниями признаков встречались в 83%. У 17% потомков имелись новые комбинации признаков (серая окраска тела и короткие крылья или черная окраска тела и нормальные крылья). Видно, что и в этих скрещиваниях проявляется склонность к сцепленному наследованию либо доминантных признаков, либо рецессивных (83%). Частичное нарушение сцепления (17% потомков) было объяснено процессом кроссинговера — обменом соответствующими участками гомологичных хромосом в профазе I мейоза (см. рис. 3.72).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
