
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Сформированная структура генеалогического дерева (рис. 5) показывает формирование башкирской популяцией самостоятельной ветви, что указывает на участие в ее создании животных иного происхождения. Высокая степень генетической консолидации башкирской популяции симментальского скота подтверждается результатами кластерного анализа: коэффициент подобия (Q) в среднем составил 84,6±2,2%. У 86,3% животных значение коэффициента Q было выше 75%. Для сравнения значения коэффициента Q в популяциях SIM_SIB, SIM_OR, SIM_A и SIM_H составили 59,1±2,4, 45,2±4,4, 74,9±2,2 и 36,3±3,2%.
Оценка информативности локусов МС для проведения экспертизы на достоверность происхождения в башкирской популяции симментальского скота показала вероятность подтверждения родителей 99,99%, вероятность исключения родителей – >99,99%.
Таким образом, сравнительные исследования пяти популяций симментальского скота (четырех чистопородных и одной голштинизированной) продемонстрировали существенный вклад башкирской популяции в общую изменчивость аллелофонда микросателлитов. Полученные данные указывают на необходимость вовлечения региональных популяций в работы по генетическому мониторингу пород крупного рогатого скота.


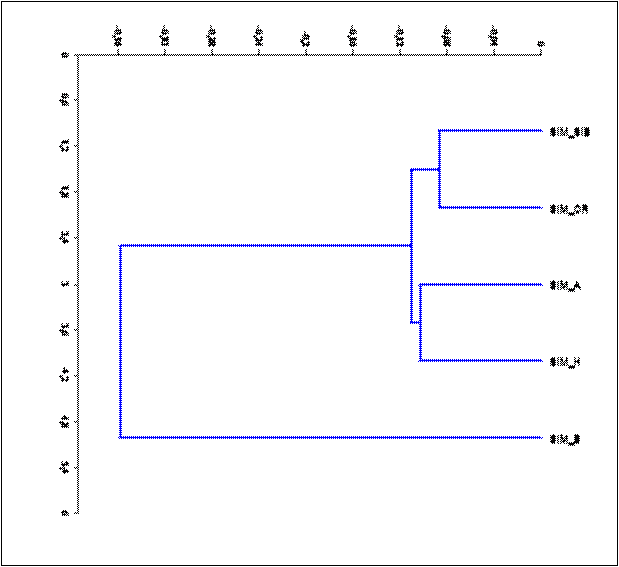
Примечание: популяции симментальского скота: сибирская (SIM_SIB), орловская (SIM_OR), австрийская (SIM_A) чистопородные и голштинизированная (SIM_H) популяции
Рис. 5. Дендрограмма филогенетического родства пяти популяций симментальского скота, построенная на основании значений генетических дистанций по Nei M. (1983) методом UMPGA
Оценку связи аллелей МС с уровнем молочной продуктивности коров проводили, сопоставляя частоты встречаемости аллелей МС с «высоким» (Xср+0,5σ) и «низким» (Xср-0,5σ) уровнем молочной продуктивности, а так же рассчитывая коэффициенты корреляции между наличием в генотипе коров определенных аллелей МС и уровнем молочной продуктивности.
3.4.1. Распределение аллелей МС в башкирских популяциях коров чёрно-пёстрой породы с различным уровнем молочной продуктивности
Анализ распределения частот встречаемости аллелей МС в группах коров чёрно-пёстрой породы CHP_AGRO с «высоким» (Xср+0,5σ) по сравнению с «низким» (Xср-0,5σ) уровнем молочной продуктивности в 1-й лактации показал наличие достоверной разницы в распределении 8 аллелей шести локусов МС. По уровню удоя - аллели 149 локуса TGLA122 (+0,088, p<0,05), 217 локуса ETH10 (+0,092, p<0,05), 248 и 256 локуса SPS,242, p<0,05 и +0,068, p<0,05, соответственно); по содержанию жира в молоке - аллели 123 локуса TGLA,033, p<0,05), 141 локуса TGLA,200, p<0,05) и 180 локуса BM1,100, p<0,05); по содержанию белка в молоке - аллель 213 локуса ETH10 (+0,135, p<0,05).
По результатам анализа 2-й лактации были выявлены 16 аллелей, частоты встречаемости которых достоверно различались между группами коров с «высоким» (Xср+0,5σ) по сравнению с «низким» (Xср-0,5σ) уровнем молочной продуктивности: по уровню удоя – аллели 234 и 236 локуса ETH,166, p<0,05 и +0,083, p<0,05, соответственно), 133 и 137 локуса BM2,111, p<0,05 и +0,084, p<0,05, соответственно), 141 локуса TGLA,118, p<0,05), 293 локуса ILST,128, p<0,05); по содержанию жира в молоке – аллели 144 локуса ETH225 (+0,091, p<0,05) и 182 локуса BM1784 (+0,093, p<0,05); по содержанию белка в молоке – аллели 181 и 183 локуса ILST,170, p<0,05 и +0,170, p<0,05), 217 локуса ETH10 (+0,098, p<0,05), 144 и 150 локуса ETH,035, p<0,05 и +0,058, p<0,05), 222 локуса ETH,074, p<0,05), 293 локуса ILST,090, p<0,05), 137 локуса BM2,229, p<0,05) и 256 локуса SPS115 (-0,070, p<0,05).
По результатам анализа популяции коров CHP_GUSP было выявлено 8 аллелей в 6 локусах МС, ассоциированных с показателями молочной продуктивности коров по 1-й лактации. По уровню удоя это аллели 81 локуса TGLA227 (+0,100, p<0,05), 222 локуса ETH185 (+0,128, p<0,05), 181 и 183 локуса ILST,047, p<0,05 и +0,047, p<0,05, соответственно); по содержанию жира в молоке – аллели 115 локуса TGLA126 (+0,274, p<0,05), 144 локуса ETH225 (+0,010, p<0,05), 81 локуса TGLA227 (-0,095, p<0,05) и 178 локуса BM1,236, p<0,05); по содержанию белка – аллели 178 и 182 локуса BM1,114, p<0,05 и +0,292, p<0,05).
Анализ данных продуктивности коров CHP_GUSP по 2-й лактации показал наличие 15 аллелей в 8 локусах, частоты встречаемости которых достоверно различались между группами коров с различным уровнем молочной продуктивности. По уровню удоя это аллели 119 и 123 локуса TGLA126 (+0,054, p<0,05 и +0,146, p<0,05), 230 локуса ETH185 (+0,199, p<0,05), 144 и 150 локуса ETH,171, p<0,05 и +0,172, p<0,05, соответственно), 127 локуса BM2113 (+0,128, p<0,05), 262 локуса BM1818 (-0,184, p<0,05); по содержанию жира в молоке – аллели 121 локуса TGLA,032, p<0,05) и 222 локуса ETH,146, p<0,05); по содержанию белка в молоке - аллели 181 и 183 локуса ILST,491, p<0,05 и +0,491, p<0,05), 117 локуса TGLA126 (+0,313, p<0,05), 183 локуса ILST005 (+0,491, p<0,05), 163 локуса TGLA122 (+0,071, p<0,05), 148 локуса ETH225 (+0,045, p<0,05) и 254 локуса SPS115 (+0,09, p<0,05).
3.4.2. Распределение аллелей МС у коров симментальской породы с различным уровнем молочной продуктивности
Анализ распределения частот встречаемости аллелей МС в группах коров симментальской породы 1-й лактации с «высоким» (Xср+0,5σ) по сравнению с «низким» (Xср-0,5σ) уровнем молочной продуктивности показал наличие достоверной разницы в распределении 5 аллелей четырех локусов МС. По уровню удоя это аллели 181 локуса ILST,100, p<0,05), 228 локуса ETH,104, p<0,05), 262 локуса BM1818 (+0,110, p<0,05), 180 и 182 локуса BM1824 (+0,237 p<0,05 и -0,289, p<0,05, соответственно); по содержанию жира в молоке – аллели 228 локуса ETH185 (+0,124, p<0,05) и 180 локуса BM1824 (+0,060 p<0,05); по содержанию белка в молоке – аллели 228 локуса ETH185 (+0,116, p<0,05), 180 и 182 локуса BM1824 (+0,081 p<0,05 и +0,052, p<0,05, соответственно). Выявленные закономерности, в большинстве случаев, сохранялись и по результатам анализа 2-й лактации.
3.5. Корреляционный анализ наличия аллелей МС и уровня молочной продуктивности коров изучаемых популяций
Результаты расчета коэффициентов корреляции между наличием в генотипе коров аллелей МС и уровнем молочной продуктивности представлены в таблице 6 (показаны только достоверные корреляции).
Как следует из данных таблицы 6, во всех исследованных нами локусах МС установлены аллели, наличие которых в генотипе коров достоверно коррелировало с показателями молочной продуктивности коров, при этом установлен породо - и популяционно-зависимый характер выявленных зависимостей.
Табл. 6. Достоверные корреляционные зависимости между аллелями МС и показателями молочной продуктивности коров
Локус | Аллель | Лактация | Коэффициенты корреляции в изучаемых популяциях | ||||||||
CHP_AGRO | CHP_GUSP | SIM_B | |||||||||
Удой, кг | Жир, % | Белок, % | Удой, кг | Жир, % | Белок, % | Удой, кг | Жир, % | Белок, % | |||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
TGLA122 | 141 | 1 | - | -0,211 | - | - | - | - | - | - | - |
2 | -0,181 | - | - | - | - | - | - | - | - | ||
149 | 1 | +0,202 | - | - | - | - | - | - | - | - | |
163 | 2 | - | - | - | - | - | +0,322 | - | - | - | |
ETH10 | 217 | 1 | +0,182 | - | - | - | - | - | - | - | - |
ETH225 | 144 | 1 | - | - | - | - | +0,211 | - | - | - | - |
2 | - | +0,201 | -0,192 | -0,212 | - | - | - | - | - | ||
148 | 2 | - | - | - | - | - | +0,222 | - | - | - | |
150 | 1 | -0,181 | - | - | - | - | - | - | - | - | |
2 | - | - | +0,171 | +0,241 | - | - | - | - | - |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
SPS115 | 248 | 1 | -0,242 | - | - | - | - | - | - | - | - |
254 | 2 | - | - | -0,182 | - | - | +0,211 | - | - | - | |
256 | 1 | +0,211 | - | - | - | - | - | - | - | - | |
TGLA126 | 115 | 1 | - | - | - | - | +0,212 | - | - | - | - |
117 | 1 | - | - | - | - | - | +0,321 | - | - | - | |
119 | 2 | - | - | - | 0,212 | - | - | - | - | - | |
121 | 2 | - | - | - | - | -0,182 | - | - | - | - | |
123 | 1 | - | -0,201 | - | - | - | - | - | - | - | |
2 | - | - | - | -0,182 | - | - | - | - | - | ||
BM1824 | 178 | 1 | - | - | - | - | +0,202 | +0,212 | - | - | - |
180 | 1 | - | -0,172 | - | - | - | - | +0,212 | +0,232 | +0,212 | |
182 | 1 | - | - | - | - | - | +0,272 | -0,322 | -0,342 | -0,312 | |
2 | - | +0,181 | - | - | - | - | - | - | - | ||
ETH185 | 228 | 1 | - | - | - | - | - | - | -0,252 | -0,252 | -0,202 |
230 | 2 | - | - | - | 0,181 | - | - | - | - | - | |
234 | 2 | -0,172 | - | - | - | - | - | - | - | - | |
236 | 2 | +0,181 | - | - | - | - | - | - | - | - | |
222 | 1 | - | - | - | +0,212 | - | - | - | - | - | |
2 | - | - | -0,252 | - | -0,191 | - | - | - | - | ||
ILST006 | 293 | 2 | -0,212 | - | -0,252 | - | - | - | - | - | - |
BM2113 | 127 | 2 | - | - | - | +0,221 | - | - | - | - | - |
133 | 2 | -0,352 | - | - | - | - | - | - | - | - | |
137 | 2 | +0,162 | - | +0,162 | - | - | - | - | - | - | |
ILST005 | 181 | 1 |
|
| -0,111 | - | -0,192 | -0,221 | -0,271 | -0,231 | |
2 | - | - | -0,172 | - | - | - | - | - | - | ||
183 | 1 |
|
| +0,151 | - | -0,192 | - | - | - | ||
2 | - | - | -0,181 | - | - | - | - | - | - | ||
TGLA227 | 81 | 1 | - | - | - | +0,182 | -0,181 | - | - | - | - |
BM1818 | 262 | 2 | - | - | - | +0,282 | - | - | +0,242 | +0,242 | +0,232 |
Примечание: 1p<0,01, 2p<0,001, «-» – не достоверно.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
