
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Г л а в а 3
Роль нервной системы в регуляции движений
7
В предыдущей главе мы рассмотрели, как мышцы, развивая усилие, воздействуют на кости, к которым они прикреплены, производя таким образом движение. Этот процесс невозможен без участия нервной системы. Как скелет остается неподвижным до тех пор, пока мышцы не приложат усилие, так и мышцы не могут сокращаться до тех пор, пока их не возбуждает нервная система. Нервная система планирует, начинает и координирует все движения человека. Эта роль нервной системы — контроль движений человека — предмет нашего изучения в данной главе.
Нервная система влияет на всю физиологическую деятельность организма человека. Нервы образуют своеобразную сеть, по которой электрические импульсы передаются практически во все участки тела, а также принимаются из них. Головной мозг действует как компьютер, интегрируя поступающую информацию, подыскивая нужный ответ и затем инструктируя соответствующие части тела, как поступить. Таким образом, нервная система обеспечивает коммуникацию и координацию взаимодействий между всеми тканями организма, а также с внешним миром.
В этой главе мы рассмотрим, как осуществляется нервный контроль произвольного движения. При этом следует помнить, что практически любая физиологическая функция, влияющая на спортивную деятельность, в той или иной мере регулируется и контролируется нервной системой. Мы рассмотрим лишь часть многогранной деятельности нервной системы в осуществлении мышечной деятельности.
СТРУКТУРА И ФУНКЦИИ НЕРВНОЙ СИСТЕМЫ
Нервная система — одна из наиболее сложных систем организма человека. Многие из ее функций еще недостаточно изучены. Поскольку данный учебник посвящен сугубо спортивной и мышечной деятельности человека, мы не будем вдаваться в ненужные подробности. Начнем с краткого ознакомления с нервной системой, затем пе
рейдем к изучению вопросов, непосредственно связанных со спортивной и мышечной деятельностью. Мы начнем с изучения основных единиц нервной системы — нейронов.
НЕЙРОН
Отдельные нервные клетки и их отростки, изображенные на рис. 3.1, называются нейронами. Обычный нейрон состоит из тела, или сомы, дендритов и аксона.
Тело содержит ядро. От тела отходят отростки — дендриты и аксон. Сбоку по направлению к
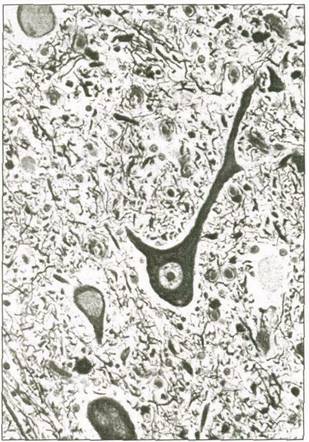
Рис. 3.1. Строение нейронов
42
аксону клетка сужается, образуя аксонный холмик, который играет важную роль в передаче импульсов.
Нейроны содержат множество дендритов. Это — рецепторы нейронов. Большинство импульсов, поступающих в нерв из соседних нейронов, как правило, поступают в нейрон через дендриты и затем передаются телу нейрона. Наряду с этим нейроны, как правило, имеют лишь один аксон. Это —нейромедиатор нейрона, передающий импульсы из тела клетки. Аксон разветвляется, образуя окончания, или терминальные фибриллы. Окончания аксона расширяются, образуя крошечные выпуклости, так называемые синаптические холмики. В них содержится множество пузырьков (мешочков), наполненных химическими соединениями — нейромедиаторами, обеспечивающими связь нейронов друг с другом (речь об этом пойдет далее).
НЕРВНЫЙ ИМПУЛЬС
Нервный импульс — электрический заряд — представляет собой сигнал, переходящий от одного нейрона к другому, пока не достигнет конечного органа, например, группы мышечных волокон, или не вернется назад в центральную нервную систему (ЦНС). Прохождение нервного импульса по нейрону можно сравнить с прохождением электрического тока по проводам. Теперь выясним, как образуется этот электрический импульс и как он проходит по нейрону.
МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ
Мембранный потенциал покоя нейрона составляет примерно —70 мВ. Это значит, что, если бы вы вставили в клетку датчик вольтметра, то увидели бы, что электрический заряд внутри клетки отличается от электрического заряда за ее пределами на 70 мВ, причем внутри клетки заряд отрицательный, а за ее пределами — положительный. Эта разница потенциалов называется мембранным потенциалом покоя (МПП). Он обусловлен разделением зарядов мембраной. При различии между зарядами мембрана поляризована.
Внутри нейрона содержится большое количество ионов калия (К'1'), тогда как снаружи — большое количество ионов натрия (Nа+). Это обусловлено тем, что натрий-калиевый насос выкачивает из клетки ионы натрия, накачивая в нее ионы калия. На основании этого можно было бы предположить, что по обе стороны мембраны заряд одинаков, однако натрий-калиевый насос, выкачивая из клетки 3 Nа+, накачивает в нее 2 К\ Кроме того, ионы калия легче проникают через мембрану, чем ионы натрия. Чтобы достичь равновесия, К^ попадают в участок низкой концентрации, поэтому некоторые из них оказываются вне клетки. Ионы натрия не могут двигаться по
добным образом. В конце концов больше положительно заряженных ионов оказывается за пределами клетки, чем внутри ее, создавая разность потенциалов. Основная функция натрий-калиевого насоса — поддержание постоянного мембранного потенциала покоя —70 мВ.
ДЕПОЛЯРИЗАЦИЯ И ГИПЕРПОЛЯРИЗАЦИЯ
Если внутри клетки заряд становится менее отрицательным относительно заряда снаружи, разность потенциалов уменьшается и, следовательно, снижается поляризация мембраны. Когда это происходит, речь идет о деполяризации мембраны. Таким образом, деполяризация имеет место при снижении мембранного потенциала покоя менее —70 мВ, приближающегося к нулю. Обычно это происходит при изменении проницаемости мембраны для Nа+.
Имеет место и обратное. При увеличении разности зарядов по обеим сторонам мембраны поляризация ее увеличивается. Этот процесс называют гиперполяризацией.
Изменения мембранного потенциала представляют собой сигналы, которые используются для приема, передачи и интеграции информации внутри клетки и между клетками. Эти сигналы бывают двух видов: ступенчатые потенциалы и потенциалы действия. Оба они являются электрическим током, образуемым движением ионов.
Подпороговые потенциалы
Подпороговые потенциалы представляют собой локальные изменения мембранного потенциала. Это либо деполяризация, либо гиперполяризация. Мембрана имеет ионные каналы, которые, в свою очередь, имеют ионные ворота, своеобразный вход в ион. Как правило, они закрыты, предотвращая проникновение ионов. При раздражении они открываются, позволяя ионам перемещаться снаружи вовнутрь и наоборот. Это приводит к разделению заряда и изменению поляризации мембраны.
Изменение локальной окружающей среды нейрона приводит в действие ступенчатые потенциалы. В зависимости от расположения и типа нейрона ионные каналы могут открываться в ответ на передачу импульса от другого нейрона или в ответ на сенсорные стимулы, такие, как изменение химической концентрации, температуры или давления.
Как мы знаем, большинство рецепторов нейрона расположены на дендритах (хотя некоторые находятся на теле клетки), а импульс всегда передается с окончаний аксона, находящихся на противоположном конце клетки. Импульс, посылаемый нейроном, проходит почти через весь нейрон. Хотя ступенчатый потенциал может привести к деполяризации мембраны, это, как правило, ло-
43
Обычно нервные импульсы идут от денд-ритов к телу, а также от тела по всей длине аксона к его терминальным фибриллам
7 |
кальное явление, и деполяризация не охватывает весь нейрон. Чтобы импульс прошел все расстояние, должен образоваться потенциал действия.
Потенциалы действия
Быстрая и значительная деполяризация мембраны нейрона называется потенциалом действия. Его продолжительность, как правило, составляет около 1 мс. Обычно мембранный потенциал изменяется с —70 мВ на +30 мВ, затем быстро возвращается к обычному уровню покоя. Как происходит это очевидное изменение мембранного потенциала?
Порог и принцип "все или ничего". Все потенциалы действия начинаются как ступенчатые. Когда стимулирование оказывается достаточным, чтобы вызвать деполяризацию порядка 15— 20 мВ, возникает потенциал действия. Это значит, что если мембрана деполяризуется от —70 до —50 — 55 мВ, в клетке возникает потенциал действия. Минимальная деполяризация, приводящая к образованию потенциала действия, называется порогом. Деполяризация меньше пороговой величины 15 — 20 мВ не приводит к образованию потенциала действия. Например, если мембран
ный потенциал изменяется от —70 до -60 мВ, изменение составляет всего 10 мВ и не отвечает порогу, поэтому потенциал действия не возникает. Если же деполяризация достигает или превышает пороговую величину, образуется потенциал действия. Это так называемый принцип или закон "все или ничего".
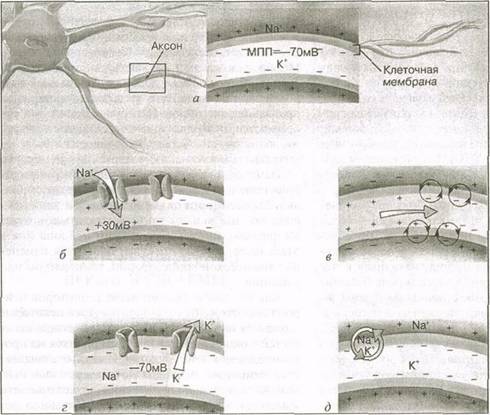
Последовательность явлений при возникновении потенциала действия. При любом возникновении потенциала действия имеет место следующая последовательность явлений (рис. 3.2):
1. Повышение проницаемости мембраны клетки для Nа+. В результате стимулирования открываются Nа+-каналы мембраны. При достижении порога проницаемость увеличивается в несколько сот раз. В клетку попадают ионы натрия. Во время этой начальной фазы количество ионов натрия, проникающих в клетку, превышает число ионов калия, выходящих из нее, вследствие чего внутренняя часть клетки становится положительно заряженной относительно внешней. Изменение напряжения (деполяризация), как правило, происходит с —70 до +30 мВ (рис. 3.2,6).
2. Снижение натриевой проницаемости. Начальная натриевая проницаемость очень кратковре-менна. Как только мембранный потенциал минует отметку "О", движение положительно заряженных ионов в клетку прекращается. Кроме того, очень быстро закрываются натриевые каналы.
3. Реполяризация. В ответ на повышение положительного заряда внутри клетки открываются калиевые каналы. Поскольку ионы калия поло-
Рис. 3.2 Последовательность явлений в возникновении потенциала действия: а — состояние покоя; б — деполяризация; в — распространение потенциала действия; г — реполяризация; д — действие Nа+/К+-насоса |
44
жительно заряжены, они перемещаются к участку с более отрицательным зарядом. Как только это происходит, снаружи клетки снова увеличивается положительный заряд, превышающий заряд внутри ее, и напряжение снова возвращается к обычному значению —70 мВ. Эта завершающая фаза — реполяризация (рис. 3.2,г).
После завершения реполяризации, прежде чем нейрон действительно вернется в свое обычное состояние покоя, должно произойти еще одно явление. Во время генерации потенциала действия Nа+ проникает в клетку. Затем для осуществления обратимости деполяризации Р^ выходит из нее. Таким образом, концентрация ионов натрия оказывается высокой внутри клетки, а концентрация К4' — снаружи ее, т. е. как раз обратной по сравнению с состоянием покоя. Чтобы окончательно поставить все на место, после завершения реполяризации активируется натрий-калиевый насос, возвращающий на свои места ионы калия и натрия (рис. 3.2,д).
Распространение потенциала действия
Выяснив, как образуется нервный импульс в виде потенциала действия, мы можем рассмотреть, как он распространяется или перемещается по нейрону. Скорость прохождения импульса через аксон зависит от двух факторов: миелиниза-ции аксона и величины нейрона.
Миелиновая оболочка. Аксоны большинства двигательных нейронов покрыты миелиновой оболочкой — жиросодержащим веществом, изолирующим клеточную мембрану. В периферической нервной системе эта оболочка образована шван-новскими клетками.
Оболочка не является цельной, участки между соседними шванновскими клетками не покрыты миелиновой оболочкой. Непокрытые участки называются перехватами Ранвье. Потенциал действия, перемещаясь по миелинизированному волокну, как будто перепрыгивает с одного перехвата на другой. Это — скачкообразная проводимость, отличающаяся более высокой интенсивностью, чем проводимость в немиелинизированных волокнах.
Скорость передачи нервного импульса по ми-елинизированньш волокнам может достигать 120 м-с~' (свыше 250 м/ч), т. е. она в 5 — 50 раз больше, чем по немиелинизированным волокнам такого же диаметра
Процесс миелинизации двигательных нейронов происходит в первые годы жизни человека, частично объясняя, почему детям необходимо какое-то время для развития координации движений. У людей, страдающих определенными неврологическими заболеваниями, например, обширным склеро
зом, происходит дегенерация миелиновой оболочки, ведущая к нарушению координации движений.
Диаметр нейрона. Скорость передачи нервного импульса также определяется размером нейрона. Проводимость нервных импульсов по нейронам большего диаметра выше, чем по нейронам небольшого диаметра, поскольку сопротивление у первых местному току более низкое.
В ОБЗОРЕ...
1. Мембранный потенциал покоя нейрона —70 мВ — результат разделения ионов натрия и калия, обусловленного главным образом действием натрий-калиевого насоса в сочетании с высокой натриевой и низкой калиевой проницаемостью мембраны нейрона.
2. Любое изменение, повышающее положительный заряд мембранного потенциала, называется деполяризацией. Любое изменение, повышающее отрицательный заряд, называется гиперполяризацией. Эти изменения имеют место при открытых ионных каналах, позволяющих ионам перемещаться из одного участка в другой.
3. При деполяризации мембраны на 15—20 мВ достигается порог, результатом которого является потенциал действия. Потенциал действия не возникает, если порог не достигается.
4. Возникновение потенциала действия включает следующую последовательность явлений:
• повышение натриевой проницаемости вследствие открытия натриевых каналов;
• снижение натриевой проницаемости при их закрытии;
• открытие калиевых каналов и реполяризация.
5. В миелинизированных нейронах импульс перемещается по аксону, "перепрыгивая" между перехватами Ранвье (участки истончения оболочки миелинового нервного волокна). Этот процесс — скачкообразная проводимость — в 5 — 50 раз выше, чем проводимость в немиелинизированных волокнах такого же диаметра.
6. Нервный импульс быстрее перемещается в нейронах большего диаметра.
СИНАПС
Связь одного нейрона с другим осуществляется благодаря потенциалу действия. После возникновения потенциала действия нервный импульс проходит через весь аксон, достигая его окончаний. Как переходит нервный импульс с одного нейрона на другой?
Связь нейронов друг с другом осуществляется с помощью синапсов. Синапс —область передачи импульса с одного нейрона на другой. Наиболее характерным видом синапсов является химический синапс, который мы и рассмотрим.

Рис. 3.3. Химический синапс между двумя нейронами (показан синоптический пузырек)
Как видно из рис. 3.3, синапс между двумя нейронами включает: концевую часть аксона нейрона, несущего импульс; рецепторы другого нейрона и пространство между этими структурами. Нейрон, посылающий импульс через синапс, называется пресинаптическим, а окончания аксона—пресинаптическими окончаниями. Нейрон, принимающий импульс на другом конце синапса, называется постсинаптическим и имеет пост-синаптические рецепторы. Окончания аксона и постсинаптические рецепторы физически не контактируют друг с другом. Их разделяет синапти-ческая щель.
Нервный импульс может передаваться через синапс только в одном направлении: от окончания аксона пресинаптического нейрона к пост
синаптическим рецепторам, как правило, находящимся на дендритах постсинаптического нейрона. Импульсы также могут поступать непосредственно на рецепторы тела нейрона, поскольку приблизительно 5 — 20 % окончаний аксона находятся ближе к телу, чем к дендритам [2].
Пресинаптические окончания аксона содержат большое количество синаптических пузырьков, или мешочков, в которых находятся нейромедиаторы. Когда импульс доходит до пресинаптических окончаний, синаптические пузырьки реагируют на него выделением своих химикалий в синаптическую щель. Эти нейротрансмиттеры затем диффундируют через синаптическую щель к постсинаптическим рецепторам нейрона, которые их связывают. Если это происходит, значит импульс достиг нейрона и может быть передан дальше.
НЕРВНО-МЫШЕЧНОЕ СОЕДИНЕНИЕ
Если связь между нейронами осуществляется в помощью синапсов, то их связь с мышечными волокнами происходит в нервно-мышечном соединении. Нервно-мышечное соединение выполняет ту же функцию, что и синапс. Даже прокси-мальная часть нервно-мышечного соединения такая же: она начинается окончаниями аксона двигательного нейрона, которые выделяют нейротрансмиттеры в пространство между двумя клетками. Окончания аксона в нервно-мышечном соединении переходят в плоские диски — концевые пластинки.
В нервно-мышечном соединении импульс принимает мышечное волокно (рис. 3.4). В месте приближения окончаний аксона к мышечному волокну оно имеет вогнутость. Образуемая таким образом впадина называется синаптическим желобом. Пространство между нейроном и мышечным волокном разделено синаптической щелью.
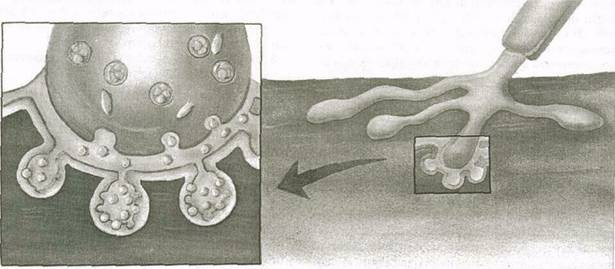
Рис. 3.4. Нервно-мышечное соединение 46
Нейромедиаторы, выделяемые окончаниями аксона, диффундируют через синаптическую щель и присоединяются к рецепторам на сарколемме мышечного волокна, мембране. Это, как правило, приводит к деполяризации, поскольку открываются каналы ионов натрия, и в мышечное волокно попадает больше натрия. Если достигается порог деполяризации, образуется потенциал действия, который распространяется через сарколемму, вызывая сокращение мышечных волокон.
Итак, мы выяснили, как передается импульс между двумя клетками. Чтобы разобраться, что происходит после передачи импульса, необходимо изучить химические сигналы, обеспечивающие передачу импульса. Итак, обратим наше внимание на нейромедиаторы.
НЕЙРОМЕДИАТОРЫ
Выявлено более 40 нейромедиаторов. Их можно разделить на быстро - и медленнодействующие, или нейропептиды (табл. 3.1). Мы рассмотрим преимущественно быстродействующие нейромедиаторы, которые обеспечивают большую часть передач нервных импульсов.
Ацетилхолин и норадреналин — основные нейромедиаторы, участвующие в регуляции физиологических реакций организма человека на физические нагрузки. Первый — основной нейро-медиатор двигательных нейронов, иннервирующих скелетную мышцу, а также многие парасимпатические нейроны. Это, как правило, возбуждающий нейромедиатор, однако он может оказывать и тормозящее действие на некоторые парасимпатические нервные окончания, например, в области сердца. Норадреналин — нейромедиатор некоторых симпатических нейронов, который также может оказывать как возбуждающее, так и тормозящее действие, в зависимости от участвующих рецепторов.
Таблица 3.1. Классификация нейромедиаторов
Быстродействующие медиаторы
Класс I Ацетилхолин
Класс II Амины: норадреналин, адреналин, допамин, серотонин и гистамин
Класс III Аминокислоты: гамма-аминомасляная кислота, глицин, глютамат и аспартат
Медленнодействующие медиаторы Гипоталамо-возбуждающие гормоны (например, тиро-тропин, выделяющий гормон соматостатин) Гипофизарные пептиды (например, р-эндорфины, ти-ротропин и вазопрессин)
Пептиды, воздействующие на мозг и кишечник (например, холецистокинин, нейротензин и лейцин-эн-кефалин)
Пептиды из других тканей (например, ангиотензин II, брадикинин и кальцитонин)
Ацетилхолин и норадреналин — основные • нейромедиаторы — химические вещества, передающие нервные импульсы через синапсы и синаптические щели
Если нейромедиатор присоединяется к пост-синаптическому рецептору, значит нервный импульс успешно передан. Затем нейромедиатор либо разрушается ферментами, либо транспортируется обратно в пресинаптические окончания для нового использования, когда поступит очередной импульс.
ПОСТСИНАПТИЧЕСКАЯ РЕАКЦИЯ

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
