
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
12.1.3. Поджелудочная железа
Поджелудочная железа (панкреас) является и пищеварительной, и эн-докринной железой. Эндокринная ткань составляет лишь 1% от веса всего органа. Она находится в железе в виде островков Лангерганса, содержащих разные клетки. Бета-клетки выделяют гормон инсулин (от лат. тзи1а - ост-ров), альфа-клетки образуют гормон глюкагон.
Инсулин повышает проницаемость мембраны мышечных и жировых клеток для глюкозы. Помогая транспорту глюкозы внутрь клетки, он способ-ствует процессам ее утилизации. Инсулин такжц имеет важное значение в от-ложении запасов углеводов в печени в виде гликогена. В частности, это важ-но в восстановительном периоде после больших физических нагрузок. При снижении уровня инсулина в крови усиливается гликогенолиз в печени. Ин-сулин стимулирует образование жира, угнетает мобилизацию его их жировых депо, способствует транспорту аминокислот и участвует в регуляции синтеза белков. Недостаточность инсулина в организме приводит к тяжелому патоло-
241
гическому состоянию (сахарной болезни, или диабету), которое характеризу-ется повышением содержания глюкозы в крови до 300-400 мг%.
Глюкагон является антагонистом инсулина. Он стимулирует расщеп-ление гликогена в печени, а также жира в жировой ткани. Секреция инсулина и глюкагона зависит от содержания глюкозы в крови. Увеличение концентра-ции глюкозы в крови повышает секрецию инсулина и прдавляет секрецию глюкагона. Секреция инсулина регулируется также вегетативной нервной системой. Раздражение блуждающего нерва усиливает выделение инсулина. Раздражение симпатических волокон уменьшает его. В то же время через симпатические нервы происходит активация секреции глюкагона.
12.1.4. Щитовидная железа
Щитовидная железа - самая крупная из эндокринных желез. Ее вес у взрослого мужчины составляет 20 г. Гормонами щитовидной железы являют-ся тироксин, трийодтиронин и калъцитонин. Тироксин и трийодтиронин различаются по количеству атомов йода в молекуле гормона, но их действие на обменные процессы качественно тождественно, различия лишь количест-венные. По-другому действует кальцитонин.
Источниками для синтеза тироксина и трийодтиронина служат ами-нокислота тирозш и йод. Полноценная функция щитовидной железы воз-можна только при достаточном содержании йода в пище.
Тироксин имеет большое значение для общего развития и роста моло-дого организма. Недостаточность функции щитовидной железы в раннем возрасте приводит к развитию так называемого кретинизма. Характерными признаками этого заболевания являются задержка роста, нарушение пропор-ций тела, задержка полового развития и умственная отсталость. У взрослого человека важнейшее значение гормонов щитовидной железы заключается в регуляции интенсивности окислительных процессов, усиливающихся под влиянием тироксина и трийодтиронина. Поэтому активность щитовидной железы отражается на уровне основного обмена, который при недостаточно-сти функции щитовидной железы уменьшается, а при патологической гипер-функции (например, при Базедовой болезни) увеличивается. Гормоны щито-видной железы - важные регуляторы термогенеза. Повышение секреции ти-роксина способствует образованию тепла в организме и, следовательно, его приспособлению к условиям воздействия холода.
Тироксин усиливает влияние адреналина и симпатической нервной системы на обменные процессы и функции сердечно-сосудистой системы. Он повышает возбудимость ЦНС, Гормоны щитовидной железы играют также важную роль в индукции синтеза белков.
Активность щитовидной железы определяется содержанием в крови тиреотропина (тиреотропного гормона, ТТГ) аденогипофиза. В свою очередь, повышенный уровень тироксина в крови угнетает продукцию этого стимуля-тора щитовидной железы. При недостатке йода в пище и питьевой воде уро-вень щитовидных гормонов в крови падает. Это обуславливает усиленную
242
секрецию ТТГ. Вследствие этого щитовидная железа гипертрофируется, хотя общая продукция гормона снижена. Это заболевание называется эндемиче-ским зобом.
Третий гормон щитовидной железы, кальцитонин, имеет важное зна-чение в регуляции обмена кальция. Основным действием капьцитонина явля-ется снижение уровня кальция в плазме крови за счет стимуляции его отло-жения в костную ткань.
12.1.5. Околощитовидные железы. Вилочковая железа. Эпифиз
Околощитовидные (паращитовидные) железы человека (обычнр их четыре) непосредственно прилегают к задней поверхности щитовидной желе-зы. Гормоном околощитовидных желез является паратгормон. Он увеличи-вает содержание кальция в крови (за счет вымывания его из костей скелета и уменьшения выведения кальция почками), воздействуя тем самым на возбу-димость и лабильность нервно-мышечного аппарата и ЦНС, а также на мно-гие секреторные процессы. Удаление паращитовидных желез приводит к клоническим судорогам, тетании и смерти. Паратгормон усиливает таюке вы-ведение фосфата почками.
Паратгормон вместе с кальцитонином регулируют содержание каль-ция в крови, которое изменяется в узких границах. Падение уровня кальция в крови усиливает активность околощитовидной железы и в результате увели-чивается содержание паратгормона в крови. Повышение содержания кальция в крови, в свою очередь, приводит к угнетению продукции паратгормона и обуславливает усиленное образование кальцитонина.
Вилочковая железа. Гормон этой железы тимозин участвует в управлении иммунологическими реакциями, а также в реализации воздейст-вия некоторых гормонов. Имеются данные, указывающие на значение вилоч-ковой железы в процессах развития организма и в приспособлении к вредным воздействиям.
Под влиянием различных сильнодействующих факторов и воздейст-вием физических нагрузок вес вилочковой железы уменьшается. Предполага-ется, что, несмотря на уменьшение веса, активность тимуса в этих случаях увеличивается.
Эпифиз. Гормон эпифиза мелатонин угнетает развитие половых же-лез, предотвращая преждевременное половое развитие, участвует также в ре-гуляции электролитного и углеводного обмена.
Секреторная активность эпифиза зависит от освещенности. Под влия-нием света продукция мелатонина уменьшается. Цикличность активности эпифиза, соответствующая смене периодов дня и ночи, представляет собой своеобразные «биологические часы» организма.
1 |
Гипофиз
Гипофиз содержит три доли - переднюю, промежуточную и заднюю.
Передняя доля гипофиза, или аденогипофиз, выполняет в эндок-ринном «оркестре» как бы роль дирижера. Такую роль он осуществляет с по-мощью тропных гормонов, регулирующих активность коры надпочечников (кортикотропина), половых желез (гонадотроптюв: фоллитропина, лютро-пина и пролактина) и щитовидной железы (тиротропина). Кроме того, аде-ногипофиз выделяет соматотропин, или гормон роста.
Под влиянием соматотропина у детей происходит ускорение роста. При недостаточной выработке этого гормона в раннем возрасте происходит задержка роста - карликовый рост. В отличие от задержки роста при крети-низме, наблюдающемся при гипофункции щитовидной железы, умственное развитие карликов нормальное. При избыточной продукции этого гормона в детском возрасте развивается гигантизм (рост человека может достигать ино-гда даже 240-250 см). Избыточная продукция соматотропного гормона после завершения роста скелета может вести к заболеванию, называемому акроме-галией. Она проявляется в непропорциональном увеличении размеров дис-тальных частей тела (пальцев, кистей, стоп, носа, нижней челюсти). Сомато-тропин действует на процессы роста непосредственно, а также через сомато-медины, которые образуются в печени под влиянием соматотропина. Сома-томедин-С стимулирует также перемещение глюкозы через клеточные мем-браны подобно действию инсулина. Поэтому его называют инсулин-подобным фактором роста.
Соматотропин повышает синтез белков, способствует транспорту аминокислот в клетку, усиливает освобождение жирных кислот из жировой ткани и в определенных условиях угнетает использование углеводов тканя-ми.
Факторами, стимулирующими или угнетающими продукцию и выде-ление тропных гормонов передней доли гипофиза, являются специфические нейрогормоны (либерины и статины), выделяемые нейросекреторными клет-ками гипоталамуса (табл. 12.1). Нейрогормоны являются факторами, посред-ством которых ЦНС участвует в регуляции эндокринных желез. Продукция тропных гормонов регулируется также содержанием гормона, образующегося под их влиянием (рис. 12.5).
В передней доле гипофиза, а также в ряде клеток ЦНС и некоторых других тканей образуются две группы нейропептидов - эндорфшы и энкефа-лты. Они способны понижать болевую чувствительность, модулировать функции ЦНС, участвовать в регуляции некоторых функций и изменять пси-хическую настройку. Эндорфин имеет важное значение в появлении чувства удовлетворения, радости и повышенного настроения (эйфории).
244 |
Таблица 12.1 |
Нейрогормоны гипоталамуса и их действие
Нейрогормоны | Действие |
Кортиколиберин | Усиливает секрецию кортикотропина |
Тиролиберин | Усиливает секрецию тиротропина |
Фоллиберин | Усиливает секрецию фоллитропина |
Люберин | Усиливает секрецию лютропина |
Соматолиберин | Усиливает секрецию соматотропина |
Соматостатин | Подавляет секрецию соматотропина |
Пролактостатин | Подавляет секрецию пролактина |
Пролактолиберин | Усиливает секрецию пролактина |
Меланолиберин | Усиливает секрецию меланотропина |
Меланостатин | Подавляет секрецию меланотропина |
Гипоталамические нейросекреторные клетки - |
ННЯСчП |
I Нейрогормоны (рилизинг
I факторы)
Обратное ингибирующее действие |
Передняя доля гипофиза |
Тропины |
Периферические железы |
Гормоны |
ТКАНИ
Рис. 12.5. Схема обратной связи: ингибирующий эффект, оказывае-мый гормонами на образование агентов, стимулирующих их секрецию
245
Промежуточная доля гипофиза у человека развита слабо. Ее продукт - меланотропин способствует образованию коричневого пигмента, который обуславливает потемнение окраски кожи.
Задняя доля гипофиза, или нейрогипофиз, выделяет два нейрогор-мона, местом образования которых являются нейросекреторные клетки гипо-таламуса. Один из них - вазопрессин - регулирует содержание воды в орга-низме и объем жидкости в кровеносных сосудах. При обеднении организма водой, т. е. при повышении осмотического давления крови, секреция этого гормона усиливается. Основное действие вазопрессина осуществляется на уровне почечных канальцев, в которых он усиливает реабсорбцию воды и тем самым уменыиает образование мочи.
Другой гормон задней доли гипофиза - окситоцин - способствует со-кращению мускулатуры матки во время родов. Во время лактации он способ-ствует отделению молока, повышая тонус гладких мышц соска.
12.2. Значение и место гормонов в регуляции обмена веществ
Обмен веществ основывается на биохимических реакциях, осуществ-ляемых при катализирующем действии ферментов. Только в узких границах условий внутренней среды ферменты обладают достаточной каталитической активностью. Эти необходимые условия (температура, рН, осмотическое дав-ление, ионный состав, напряжение кислорода, содержание воды) составляют жесткие гомеостатические константы организма. Этим жестким парамет-рам присущ весьма ограниченный диапазон между уррвнем константного состояния и отклонением, не совместимьш с жизнью. Наряду с жесткими константами внутренняя среда характеризуется также пластическими кон-стантами, обладающими широким диапазоном изменчивости. Всякое измене-ние условий существования организма и любая активация процессов жизне-деятельности создает угрозу смещения констант внутренней среды. Чтобы исключить это, скомпенсировать смещающее воздействие и восстановить константное состояние, включаются приспособительные реакции. Совокуп-ность скоординированных физиологических функций, направленных на обеспечение постоянства жестких констант внутренней среды организма, обозначается термином «гомеостатическая регуляция». При этом широко используется изменчивость пластических констант внутренней среды.
Гомеостатической регуляцией обеспечивается необходимость в гиб-кой регуляции обменных процессов. Однако необходимость в регуляции об-менных процессов обуславливается также выполнением различных актов жизнедеятельности.
Регуляция обмена веществ. Главный путь управления обменом ве-ществ - изменение активности ферментов. Активность ферментов изменя-ется в результате обратимых изменений в структуре молекул фермента, а
246
также в результате увеличения или уменьшения количества молекул фермен-та. Как было отмечено, активность ферментов зависит от условий среды. Кроме этого важно присутствие коферментов. Во многих случаях в их роли выступают витамты.
Уровнями регуляции обменных процессов являются: 1) клеточная ав-торегуляция; 2) гормональная регуляция; 3) нервная регуляция.
Клеточная авторегуляция основывается на противоположных влия-ниях субстратов (С) и продуктов (П) на ферменты, катализирующие биохи-мические реакции. Если биохимическую реакцию представить как:
Ф,
С :<=* П, ф2
то фермент Фь катализирующий превращение субстрата в продукт, активи-руется высоким содержанием субстрата и низким содержанием продукта. В то же время фермент Ф^, катализирующий обратное направление реакции (продукт превращается обратно в субстрат), ингибируется этими же воздей-ствиями. Если содержание продукта начинает превышать содержание суб-страта, то происходят противоположные изменения: активность фермента Ф) ингибируется, а активность фермента Ф2 повышается, вследствие этого оста-навливается реакция в первоначальном направлении и заменяется реакцией в противоположном направлении.
Биохимические реакции составляют целостные целенаправленные системы, поэтому регулируется не каждая отдельная реакция через катализи-рующий ее фермент, а система (цепь или цикл) реакций в целом.
Основной целью авторегуляции является обеспечение постоянства, биохимической среды внутри клеток, в частности - отношение между суб - ; стратами и продуктами различных реакций. Однако клеточная авторегуляция не удовлетворяется, если необходима значительная мобилизация ресурсов организма. В частности, при выполнении спортивных упражнений или дру - ; гих форм напряженной мышечной работы необходимо продолжительное и интенсивное протекание биохимических реакций в одном направлении. Так, , пробегание 400-метровой дистанции с соревновательной скоростью возмож - ' но только тогда, когда гликогенолиз в мышцах не ингибируется по мере на - 1 копления лактата. Таким образом, для обеспечения достаточно эффективной | мобилизации ресурсов необходимо вмешательство в клеточную авторегуля - | цию. Это становится возможным через гормональную и нервную регуляцию. !
Гормональная регуляция осуществляется через двоякие воздействия на | активность ферментов. Одни гормоны обуславливают изменения молекул \ фермента, превращающие менее активные формы фермента в активные, или! наоборот. Другие гормоны оказывают непосредственное влияние на синтез и ' распад ферментов, изменяя тем самым количество молекул фермента. Благо-даря обоим изменениям возможно поддержание биохимических реакций в одном направлении, несмотря на снижение содержания субстрата и накопле-ние продукта. Например, значительная мобилизация запасов гликогена не - |
г
I
247
смотря на накопление лактата при беге на 400 м становится возможной бла-годаря действию адреналина на активность гликоген-фосфорилазы в рабо-тающих мышцах.
Важным фактором, регулирующим интенсивность образования и сек-реции гормонов, является характер регулируемых ими проЦессов. Как только изменения, вызываемые каким-либо гормоном, достигают определенной ве-личины, образование и выделение этого гормона уменьшается. В ряде случа-ев увеличивается продукция другого гормона, регулирующего противопо-ложно направленный процесс. Таким образом, высокие содержания в крови продуктов обменных процессов, образующихся под влиянием гормона, не-редко подавляют активность соответствующей железы, а низкое содержание этих продуктов может быть стимулом активации ее деятельности (первый вариант механизма отрицательной обратной связи).
Щитовидная железа, кора надпочечников и половые железы регули-руются тропными гормонами передней доли гипофиза. В этом взаимодейст-вии важную роль играет основное влияние уровня гормонов щитовидной же-лезы, коры надпочечников и половых желез на продукцию соответствующих тропных гормонов (см. рис. 12.5). Высокий уровень концентрации гормонов указанных желез в крови подавляет, а низкий уровень - усиливает секрецию тропных гормонов (второй вариант механизма отрицательной обратной свя-зи).
Таким образом, гормональная регуляция также связана с механизма-ми, ограничивающими значительную мобилизацию ресурсов организма. По-этому контроль продукции гормонов необходим со стороны ЦНС.
Нервная регуляция обеспечивает взаимодействие между разными тка-нями, органами и частями организма, а также между организмом и внешней средой. Нервная регуляция обменных процессов осуществляется путем нерв-ных влияний на интенсивность продукции гормонов в железах внутренней секреции, т. е. управлением гормональной регуляцией. Обнаружена также возможность непосредственных нервных влияний на активность ферментов и условия протекания биохимических реакций. Это обеспечивается влиянием медиаторных веществ, освобождающихся из периферических синапсов.
Существуют два канала влияния ЦНС на эндокринные функции. Одни эндокринные железы активируются через вегетативные нервы, иннервирую-щие эндокринные железы, другие - двухэтапным механизмом. Первый этап заключается в образовании тйрогормонов нейросекреторными клетками промежуточного мозга (гипоталамуса), которые управляют секрецией троп-ных гормонов аденогипофиза. Вторым этапом является стимуляция активно-сти периферических эндокринных желез тропными гормонами аденогипофи-за(рис. 12.6).
248 |
|
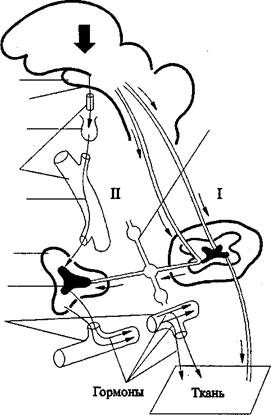
Головной мозг |
Гипоталамус Нейросекрет Гипофиз Кровеносные сосуды Кортикотропин |
Вегетативный нерв |
Спинной мозг |
Кора надпочечников Мозговой слой надпочечников Кровеносные сосуды |
Нерв |
I |
Рис. 12.6. Два канала передачи воздействий центральной нервной
системы к периферичвским органам и тканям: прямой нейрональный
(I) и нейрогормональный (II)
I. Нервные импульсы, возникающие на разных уровнях центральной нервной системы, направляются по периферическим нервам к органам и тканям. II. Возбуждение, возникающее в разных отделах головного мозга, оказывает влияние на периферические органы и ткани через эндокринные железы, т. е. путем гормональной регуляции. Этим управляет гипоталамус, который пере-дает влияния центрапьной нервной системы на эндокринные железы по двум путям: 1) непосредственно через вегетативные нервы, доходящие до эндок-ринных желез, 2) нейросекреторным путем - клетки гипоталамуса, обладаю-щие нейросекреторной активностью, выделяют специфические нейрогормоны, которые через кровоток поступают в аденогипофиз, изменяя интенсивность секреции его тропных гормонов. Последние доходят через кровообращение к эндокринным железам, усиливая продукцию соответствующих гормонов. Кровь переносит гормоны к органам и тканям, подвергаемым их регуляторно-му влиянию.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
