
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ры, или посредники. Ими могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных узлах), норадреналин (в окончаниях симпа-тических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Приходящие к окончанию аксона нервные импульсы вызывают выход ме-диатора из синаптических пузырьков и выведение его в синаптическую щель.
По характеру воздействия на последующую нервную клетку разли-чают возбуждающие и тормозящие синапсы. В возбуждающих синапсах ме-диаторы (например, ацетилхолин) связываются со специфическими макро-молекулами постсинаптической мембраны и вызывают ее деполяризацию. При этом регистрируется небольшое и кратковременное (около 1 мс) коле-бание мембранного потенциала в сторону деполяризации, или возбуждаю-щий постсинаптический потенциал (ВПСП). Для возбуждения нейрона необ-ходимо, чтобы ВПСП достиг порогового уровня. Для этого величина депо-ляризационного сдвига мембранного потенциала должна составлять не менее 10 мВ. Действие медиатора очень кратковременно (1-2 мс), после чего он расщепляется на неэффективные компоненты (например, ацетилхолин рас-щепляется ферментом холинэстеразой на холин и уксусную кйслоту) или поглощается обратно пресинаптическими окончаниями (например, норадре-налин).
В тормозящих синапсах содержатся тормозные медиаторы (например, гамма-аминомасляная кислота). Их действие на постсинаптическую мембра-ну вызывает усиление выхода ионов калия из клетки и увеличение поляриза-ции мембраны. При этом регистрируется кратковременное колебание мем-бранного потенциала в сторону гиперполяризации - тормозящий постсинап-тический потенциал (ТПСП). В результате нервная клетка оказывается за-блокированной. Возбудить ее труднее, чем в исходном состоянии. Для этого
16
понадобится более сильное раздражение, чтобы достичь критического уров-ня деполяризации.
Возникновение импульсного ответа нейрона. На мембране тела и дендритов нервной клетки находятся как возбуждающие, так и тормозящие синапсы. В отдельные моменты времени часть их может быть неактивной, а другая часть оказывает активное влияние на прилегающие к ним участки мембраны. Общее изменение мембранного потенциала нейрона является ре-зультатом сложного взаимодействия (интеграции) местных ВПСП и ТПСП всех многочисленных активированных синапсов. При одновременном влия-нии как возбуждающих, так и тормозящих синапсов происходит алгебраиче-ское суммирование (т. е. взаимное вычитание) их эффектов. При этом возбу-ждение нейрона возникает лишь в том случае, если сумма возбуждающих постсинаптических потенциалов окажется больше суммы тормозящих. Это превышение должно составлять определенную пороговую величину (около 10 мВ). Только в этом случае появляется потенциал действия клетки. В це-лом возбудимость нейрона зависит от его размеров: чем меныие клетка, тем выше ее возбудимость.
С появлением потенциала действия начинается процесс проведения нервного импульса по аксону и передача его на следующий нейрон или ра-бочий орган, т. е. осуществляется эффекторная функция нейрона. Нервный импульс - основное средство связи между нейронами.
Таким образом, передача информации в нервной системе происходит с помощью двух механизмов - электрического (ВПСП, ТПСП, потенциал действия) и химического (медиаторы).
1.2.3. Особенности деятельности нервных центров
Свойства нервных центров в значительной мере связаны с особенно-стями проведения нервных импульсов через синапсы, связывающие различ-ные нервные клетки.
Проведение возбуждения через нервные центры. Нервным цен-тром называют совокупность нервных клеток, расположенных в определен-ном отделе ЦНС и осуществляющих какую-либо функцию. Эти центры от-вечают соответствующими рефлекторными реакциями на внешнее раздра-жение, поступившее от связанных с ними рецепторов. Клетки нервных цен-тров реагируют и на непосредственное их раздражение веществами, находя-щимися в протекающей через них крови (гуморальные влияния).
Проведение волны возбуждения от одного нейрона к другому через синапс происходит химическим путем - с помощью медиатора. Медиатор содержится лишь в пресинаптической части синапса и отсутствует в постси-наптической мембране. Поэтому важной особенностью проведения возбуж-дения через нервные центры является одностороннее проведение нервных влияний через синапс, которое врзможно лишь от пресинаптической мем-
17 |
браны к постсинаптической и невозможно в обратном направлении. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление от афферентных нейронов к вставочным и затем к эфферент-ным - мотонейронам или вегетативным нейронам.
Болыиое значение в деятельности нервной системы имеет другая осо-бенность проведения возбуждения через нервные центры - замедленное про-ведение через синапс. Замедление связано с затратой времени на процессы, происходящие от момента подхода нервного импульса к пресинаптической мембране до появления в постсинаптической мембране возбуждающих или тормозящих потенциалов. Этот интервал называется синаптической задерж-кой. В болыиинстве центральных нейронов он составляет около 0,3 мс. По-сле этого требуется еще время на развитие ВПСП и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс зани-мает примерно 1,5 мс. При утомлении, охлаждении и ряде других воздейст-вий длительность синаптической задержки возрастает. Если же для осущест-вления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.
При рефлекторной деятельности общее время от момента нанесения внешнего раздражения до появления ответной реакции организма - так на-зываемое скрытое, или латентное, время рефлекса - определяется в основном длительностью проведения возбуждения через синапсы. Величина латентно-го времени рефлекса служит важным показателем функционального состоя-ния нервных центров. Измерение латентного времени простой двигательной реакции человека на внешний сигнал широко используется в практике для оценки функционального состояния ЦНС (рис. 1.6).
|
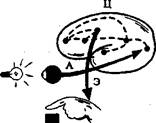
Рис. 1.6. Определение времени
простой двигательной реакции
человека
А - афферентный путь, Э - эффе-рентный путь, Ц — центральные пути рефлекторной реакции. На кривой внизу: С - отметка вспышки света, О - отметка нажима на кнопку, 1=150 мс - латентное время реакции
150 мс |
о |
18 |
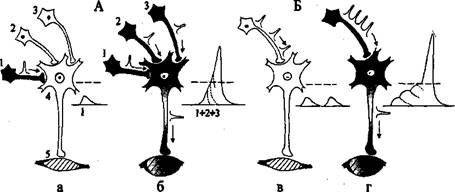
Суммация возбуждения. В ответ на одиночные афферентные им-пульсы, идущие от рецепторов к нейронам, в пресинаптической части синап-са освобождается небольшое количество медиаторов. При этом в постсинап-тической мембране нейрона обычно возникает ВПСП - неболыпая местная деполяризация. Для того чтобы общая по всей мембране величина ВПСП достигла порога возникновения потенциала действия, требуется суммация на мембране клетки многих подпороговых ВПСГЪ Лишь в результате такой суммации возбуждения возникает ответ нейрона. Различают пространствен-ную и временную суммацию (рис. 1.7).
|
Потенциал действия |
Порог |
Потенциал понп |
Рис. 1.7. Пространственная (А) и временная (Б) суммация возбуждения
1, 2, 3 - нейроны различных рефлекторных дуг, 4 - мотонейрон спинного мозга, 5 - иннерви-руемая мышца; а - одиночное возбуждение отдельного нейрона (1) и подпороговый возбуж-дающий постсинаптический потенциал - ВПСП (1); б - одновременный приход возбуждения по трем пресинаптическим волокнам (1, 2 и 3), пространственная суммация ВПСП (1+2+3) на мембране мотонейрона и проведение потенциала действия к мышце (стрелка); в - подпоро-говый ВПСП при низкой частоте нервных импульсов; г - временная суммация ВПСП при высокой частоте нервных импульсов, появление потенциала действия мотонейрона и его проведение к мышце
Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пре-синаптическим волокнам. Одномоментное возбуждение синапсов в различ-ных участках мембраны нейрона повышает амплитуду суммарного ВПСП до пороговой величины. В результате возникает ответный импульс нейрона и осуществляется рефлекторная реакция. Например, для получения ответа дви-гательной клетки спинного мозга обычно требуется одновременная актива-ция 50-100 афферентных волокон от соответствующих периферических ре-цепторов.
19
Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражений. Если интервалы между поступающими импульсами достаточно коротки и ВПСП нейрона от предыдущих раздражений не успевают затухать, то последующие ВПСП накладываются друг на друга. Когда деполяризация мембраны нейрона дос-тигнет критического уровня, возникает потенциал действия. Таким образом, даже слабые раздражения через некоторое время вызывают ответные реак-ции организма.
Трансформация и усвоение ритма. Характер ответного разряда нейрона зависит не только от свойства раздражителя, но и от функциональ-ного состояния самого нейрона (его мембранного заряда, возбудимости, ла-бильности). Нервные клетки обладают свойством изменять частоту пере-дающихся импульсов, т. е. свойством трансформацшритма.
При высокой возбудимости нейрона (например, после приема кофеи-на) может возникать учащение импульсации (мультипликация ритма), а при низкой возбудимости (например, при утомлении) происходит урежение рит-ма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения потенциала действия. Эти изменения частоты импульсации могут усиливать или ослаблять ответные реакции.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
