
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
У всех тюленей новые двигательные рефлексы вырабатываются легко и закрепляются прочно, но скорость выработки реакций на различные сигналы зависит от их сложности. После длительного перерыва в занятиях угасшие рефлексы восстанавливаются достаточно быстро – необходимо 2-3 предъявления, и команда снова выполняется правильно. Тюлени предлагают большое количество различных движений, которые впоследствии можно закрепить, используя определенную команду.
В целом, сравнивая поведение морского зайца и тюленей других видов (серого, гренландского) в океанариуме, следует отметить флегматичность, медлительность при выполнении команд. Эти особенности, вероятно, связаны с образом жизни лахтака, кормящегося в природных условиях главным образом малоподвижными бентосными организмами и не являющегося, следовательно, таким активным хищником, как большинство настоящих тюленей.
Глава 4 РАСПОЗНАВАНИЕ ТЮЛЕНЯМИ ЗРИТЕЛЬНЫХ СТИМУЛОВ
4.1 Выработка дифференцировочных условных рефлексов на двумерные зрительные стимулы
На первом этапе работы тюленей обучали "новым", ранее не входившим в их репертуар на представлениях движениям, выполняемым по определенной жестовой команде. 1 – кольцо – самка серого тюленя №2 совершает полный круг вокруг экспериментатора, стоящего на помосте, самец серого тюленя №1 выполняет "колечко" (прогибается в спине, касаясь носом хвоста); 2 – треугольник – тюлень №2 выходит на левый край бассейна, тюлень №1 – "стыдно" (закрывает глаз передним ластом).
Обучение на 1 этапе заняло у животных разное количество занятий: 9 у тюленя №2 и 10 у тюленя №1. У тюленя №2 уровень правильных ответов (выполнение движений 1 и 2) составил 96.0±2.3%), у тюленя №1 – 90.3±2.8%, за последние 5 тренировок 96.3±0.9% тюленю №2 на первом этапе потребовалось 180 предъявлений, тюленю №1 – 250. Высокую скорость выработки новых условных рефлексов можно объяснить тем, что у животных сформирована установка на обучение по жестовым командам.
На 2 этапе проводилось установление и закрепление связи: зрительный стимул (кольцо или треугольник) - жест - движение животного. Данный этап занял у тюленя №1 – 16, у тюленя №2 – 20 занятий. Тюлень №2 получил 330 предъявлений (97.8±1.1% правильных ответов), тюлень №1 - 289 (99.6±0.4% правильных ответов).
В последующих опытах животным предъявляли только таблички-символы (кольцо и треугольник попеременно). В результате среднее число правильных зрительных дифференцировок (соответствие движения животного определенному изображению) после 22 опытов с самцом (тюлень №1) и 20 с самкой (тюлень №2) по 25 предъявлений в каждом незначительно превышало случайную величину 58.3±2.7% и 61.5±3.9% соответственно. При применении дополнительных методических приемов для переключения внимания животных с экспериментатора на табличку с изображением уровень правильных ответов в отдельных опытах повышался до 80-100%. При обучении по этой же стандартной методике морскому котику потребовалось 79 опытов с 51 предъявлением в каждом для достижения уровня дифференцировки 95%. В начале же обучения наблюдалось 35 – 40% межсигнальных (т. е. неправильных) реакций (Дашевский, Близнюк, 1989). Кроме того, в отличие от наших экспериментов, в опытах, которые проводили упомянутые авторы, использовались животные, не участвовавшие в представлениях.
По описанной выше схеме опыт был проведен с самцом серого тюленя №6 и самцом морского зайца (тюлень №7). У животных проводилась выработка условных рефлексов на зрительные раздражители, "крест" и "квадрат". Для установления связи "геометрическая фигура – сигнал-жест тренера – выполнение элемента – подкрепление" были проведены три занятия с комплексным раздражителем: изображение в сочетании с жестом и голосовой командой тренера. На первых 2-х занятиях с тюленем №7 и 3 занятиях с тюленем №6 уровень правильных ответов составил 100%. В последующих опытах, без сопровождения голосовой командой, было замечено, что животные сосредотачиваются на руках тренера, воспринимая любое движение за жест, а предъявляемая фигура остается без внимания. Этот эксперимент подтвердил ранее высказанное мнение (Дашевский, Близнюк, 1989), что при дифференцировке зрительных стимулов у ластоногих ведущую роль играет переключение внимания животного с экспериментатора на предъявляемый зрительный раздражитель.
4.2 Различение тюленями геометрических фигур разной степени подобия
В данной серии экспериментов методика работы по различению двумерных геометрических фигур серым тюленем и морским зайцем была изменена: с целью достижения переключения внимания животного на таблички-символы, их расположили на стенке бассейна, а не на помосте, и животные осуществляли выбор, подплыв к распознаваемым объектам.
С морским зайцем было проведено 17 занятий (рис. 1) с предъявлением ему 5 вариантов креста. В 4-х вариантах (опыты 1-13) морской заяц (тюлень №7) очень уверенно и быстро дифференцировал крест от квадрата. Однако в варианте 5 (ширина сторон креста 21 см) достоверная дифференцировка отсутствовала, из чего можно заключить, что такая фигура воспринимается животным как квадрат. При работе с вариантом 5 было отмечено, что животное использовало тактику выбора в большинстве случаев фигуры, расположенной слева, что позволяло получать подкрепление за случайно выбранный правильный ответ. В отличие от тюленя №7, тюлень №6 уверенно распознал и 5-й вариант креста, наиболее сходный с квадратом.
Вероятно, выявленные различия в способности распознавать геометрические фигуры обусловлено разным образом жизни, способом питания этих животных. Серые тюлени – активные хищники, а морские зайцы бентофаги и при поиске пищи для них большее значение имеет чувствительность вибрисс, чем зрение. Суммируя вышесказанное, можно сделать вывод, что настоящие тюлени хорошо дифференцируют плоские геометрические фигуры по форме, а не только объемные, как считалось ранее.
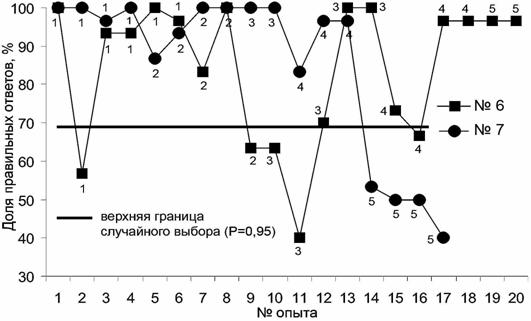
Рис. 1. Различение квадрата и креста самцом морского зайца (тюлень №7) и самцом серого тюленя (тюлень №6), 1-5 – варианты креста
4.3 Распознавание трехмерных зрительных раздражителей
Выработка дифференцировочных условных рефлексов на трехмерные зрительные раздражители
Работа проводилась на двух молодых серых тюленях самце и самке (тюлень №3 и тюлень №4). К началу эксперимента животные имели навыки выполнения различных движений по жестовым командам. Всего с тюленем №3 проведено 24 опыта с числом предъявлений 32 в каждом, с тюленем №4 – 24 (число предъявлений 40).
Эксперименты начали с установления и закрепления связи: предъявление предмета – сигнал-жест экспериментатора – выполнение адекватного движения – подкрепление. Соответствие стимулов и движений было следующим: 1 - тор (кольцо) – кружиться в воде вокруг своей оси; 2 – шар (мячик) – показать зубы; 3 – цилиндр (отрезок трубы) – протянуть из воды правый передний ласт.
Экспериментатор показывал животному предмет, делал определенный жест, после чего тюлень должен был выполнять соответствующее движение. Затем (при правильном ответе) следовало подкрепление. Занятия проводились с тюленем №3 в апреле, а с тюленем №4 в сентябре. Это явно сказалось на результатах опытов, поскольку она(самка серого тюленя) имела возможность наблюдать за занятиями с самцом. В ответ на предъявление кольца во время одной из тренировок при подготовке к выступлению она сразу, без обучения, выполнила движение, которое соответствует предъявлению кольца у самца. В данном случае ярко проявилась способность тюленя к обучению при помощи подражания.
Данный этап работы занял у тюленя №3 и у тюленя №4 по 6 занятий. Тюлень №4 получила 220 предъявлений (79.93±5.08% правильных ответов), тюлень №3 – 161 (71.68±7.32% правильных ответов). В последующих опытах по выработке дифференцировки попеременно предъявляли по одному из трех предметов (кольцо, мячик и трубу). Порядок предъявления стимулов был псевдослучайным (один стимул не более трех раз подряд). Таких занятий проведено по 18 с обоими тюленями. Среднее число правильных ответов у тюленя №3 составило 92.87±1.57%, (182 предъявления мяча, 188 – кольца, 224 – трубы), у тюленя №4 – 81.78±2,84% (181, 204, 383 предъявления, соответственно) (рис. 2).
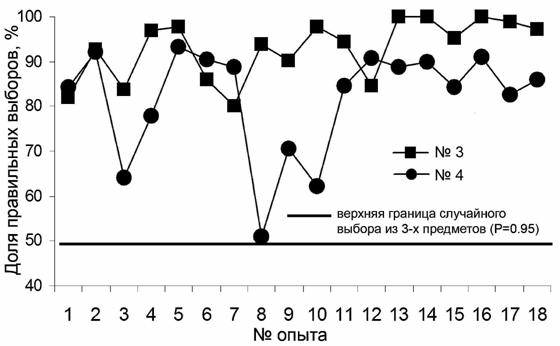
Рис. 2. Динамика выработки дифференцировки трехмерных зрительных раздражителей
Уровень правильных дифференцировок у тюленя №3 был выше, чем у тюленя №4. В отличие от самца(тюлень №3), самка (тюлень №4) в течение 6 месяцев, предшествовавших экспериментам, выполняла движения по жестовым командам во время представлений в океанариуме. Это, по-видимому, и обусловило более низкий уровень дифференцировок. Результаты же самца свидетельствуют о том, что обучение распознаванию трехмерных зрительных раздражителей у этого тюленя достигнуто. Уже в первых опытах (№4 и №5) правильная дифференцировка наблюдалась более чем в 95% случаев, а в опытах №№ 13, 14 и 16 её уровень был близок к 100 процентам. Тем не менее, обучение самки также можно считать успешным: начиная со второй половины серии экспериментов и до их окончания результаты самки отличались высокой стабильностью (рис. 2, опыты №№ 11-18).
4.4 Выбор предметов по образцу
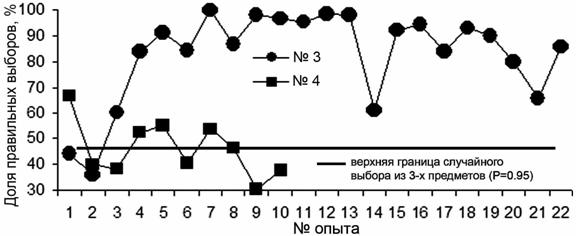
После обучения правилу выбора по образцу, тюлени из трех закрепленных на стенке вольера предметов выбирали, прикасаясь носом, один, соответствующий предъявленному образцу. Таких опытов с самкой (тюлень №4) проведено 10 (575 предъявлений, 61.48±5.79% правильных ответов), с самцом (тюлень №3) 22 (1135 предъявлений, 82.8±3.8% правильных ответов) (в опытах с 9-го по 13-й – не ниже 95%) (рис. 3).
При работе с самкой отмечено, что она не обозначает предмет, который висит дальше всех от помоста. В результате выбор осуществляется между двумя предметами. Если "дальний" из них именно тот, который соответствует предъявленному образцу, то выбор будет заведомо неправильным. Кроме того, самец был постоянно занят только в экспериментальной работе, самка участвовала в ней в то время, когда была свободна от представлений. На представлениях закреплялось и поддерживалось на высоком уровне выполнение рефлексов на сигналы-жесты.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
