
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рис. 3. Множественный выбор по образцу
Предъявление предметов в предыдущих экспериментах осуществлялось "псевдослучайно". Это могло привести к выработке у животных рефлексов не на присутствие определенных предметов в поле зрения, а на интервалы времени их предъявления или другие параметры. Поэтому заключительная серия экспериментов была проведена с использованием таблицы случайной комбинации трех чисел (рис. 4). Всего с самцом (тюлень №3) проведено 21 занятие со средним числом предъявлений 36. Тюлень получил 754 предъявления (258 – мяч, 264 – кольцо, 232 - труба). Среднее число правильных ответов составило (86.76±1.52%). В конце экспериментов (опыты 18 и 19) оно превысило 95%. Таким образом, и при псевдослучайном, и при случайном предъявлении предметов "выбор по образцу" осуществлялся с достаточной достоверностью.
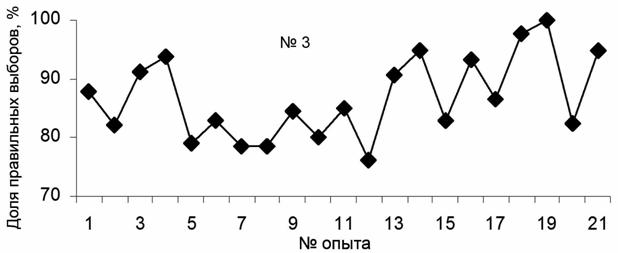
Рис. 4. Динамика обучения тюленя "выбору по образцу" из множества
Глава 5 РАСПОЗНАВАНИЕ ТЮЛЕНЯМИ СЛУХОВЫХ СТИМУЛОВ
5.1 Дифференцировка звуков различной частоты морским зайцем
В результате проведенных экспериментов морской заяц (тюлень №7) был обучен производить определенные движения в ответ на звуковые сигналы различной частоты. Звуки были выбраны в диапазоне вокализации морских зайцев: от 01.01.01Гц (Свиженко, 1983). Для достижения 100%-й точности выполнения первого из вновь выработанных рефлексов потребовалось 244 предъявления раздражиГц – "ладушки"), второго – 70 (1400 Гц – лежать), третьего – 111 (200 Гц – "стыдно"). Ускорение обучения реакции на второй сигнал более чем в 3 раза вполне объяснимо формированием "установки на обучение".
При предъявлении морскому зайцу звуковых раздражителей осуществлялась не только выработка новых условных рефлексов, но и дифференцировка раздражителей. На этапах эксперимента, когда вводили новый звук, происходила конкуренция между двумя, а затем и между тремя условными сигналами. Это выглядело как осуществление тюленем выбора – каким движением следует ответить и, вероятно, значительно повлияло на уровень правильных реакций. Средний уровень дифференцировки двух звуков составил 90.2±4.4%, трёх – 75.2±4.5%. Результат экспериментов, проведенных после четырёхмесячного перерыва, оказался в значительной степени неожиданным: сначала тюлень не вполне уверенно дифференцировал все 3 сигнала (опыт № 28, 200 Гц – 63%, 700 Гц – 75%, 1400 Гц – 100%), а затем (опыт № 29) продемонстрировал 100%-е их распознавание (рис. 5).
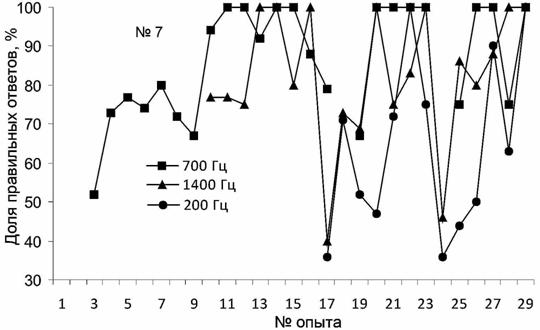
Рис. 5. Динамика формирования у морского зайца дифференцировки звуков различной частоты
5.2 Определение минимального частотного интервала между двумя разными звуковыми сигналами, распознаваемыми тюленями
5.2.1 Морской заяц
Первая пара звуков (200 Гц и 700 Гц), предложена тюленю для распознавания, поскольку ранее проведенные эксперименты позволяют нам, во-первых, не тратить время на выработку новых двигательных навыков, которые животное должно выполнять при предъявлении звукового раздражителя; а во-вторых, эту пару звуков морской заяц способен надежно дифференцировать.
Постепенно (опыты с 18 по 33) (рис. 6), тюлень стал распознавать звуки по относительному признаку "выше – ниже" уже при первом предъявлении новых пар звуков, что может свидетельствовать о высоком уровне установки на обучение и способности тюленя дифференцировать звуки не только по абсолютному, но и по относительному признаку.
Уменьшение числа правильных реакций на звуки 420 Гц и 480 Гц можно объяснить возможным отклонением в здоровье животного. Подобные спады, вплоть до отказа работать, встречаются при обучении всех тюленей в океанариуме. Определить причины этого, как правило, бывает трудно, за исключением случаев, когда явно присутствует отвлекающий фактор (например, сильный необычный запах, шум и т. п.), симптомы заболевания. Снижение пищевой мотивации наблюдается у животных, которые активно участвуют в представлениях океанариума. Если во время первого эксперимента заяц не участвовал в представлениях, то во время проведения данного эксперимента он был включен в ежедневные демонстрационные программы.
В опытах 54–59 частотный интервал между двумя основными звуками составляет 10 Гц, и животное их уверенно дифференцирует с 95%-м уровнем правильных ответов в двух опытах подряд (рис. 6, опыты 58-59). Поэтому в последующих экспериментах частотный интервал между распознаваемыми звуками уменьшали на 1 Гц.
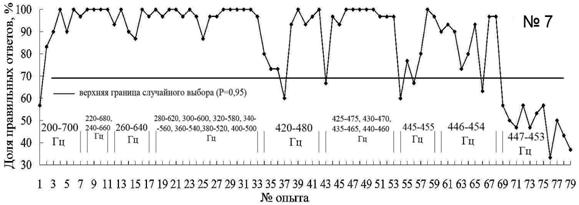
Рис. 6. Распознавание морским зайцем пар звуков разной частоты (над осью обозначены применявшиеся для дискриминации частоты пар звуковых сигналов)
В результате установлено, минимальный интервал между звуками, которые уверенно дифференцирует морской заяц, составляет 8 Гц. Результаты опытов 69–78 (рис. 6), где частотный интервал составил 6 Гц, показали, что морской заяц не дифференцирует эти звуки. При этом отмечено, что при предъявление звуковых раздражителей животное просто выполняет одно из двух движений. Во время опытов 69 – 77 (рис. 6) между предъявлениями звуковых раздражителей, животное вело себя беспокойно: двигалось по помосту, переворачивалось на спину, пытаясь получить пищевое подкрепление.
5.2.2 Гренландский тюлень
Работа с гренландским тюленем началась с выработки условных рефлексов "стыдно" и "ладушки" на голосовую команду тренера. Ранее животное было уже обучено выполнять эти действия по жестовым сигналам.
Первые 3 занятия предъявление звуков 200 и 700 Гц сопровождалось голосовой командой тренера (рис. 7). Два занятия животное уверенно дифференцировало эти две команды, на третьем занятии голосовая команда подавалась выборочно, и животное дифференцировало звуки с точностью 76.6%. С занятия №9 гренландский тюлень со 100%-ой уверенностью дифференцировал эти два звука.
В опытах 41-64 был отмечен спад правильных реакций на предъявляемые звуки. У животного наступил период линьки, что оказало существенное влияние на уровень физиологической активности и пищевой мотивации. Кроме того, сказалось влияние внешних факторов, как, например, в опыте №62 (рис. 7), когда при занятиях с гренландским тюленем присутствовали дети из детской художественной школы, которые рисовали тюленей.
При предъявлении пары звуков 425–475 Гц у гренландского тюленя, в отличие от морского зайца, не отмечено резкого снижения правильных реакций. Но также следует отметить, что животное уверенно дифференцирует пару 440 –460 Гц, тогда как при предъявлении пары 445 – 455 Гц 95%-ый уровень правильных реакций достигнут не был, а в опытах № 97, 104 и 105 составил 53.33% (рис. 7).
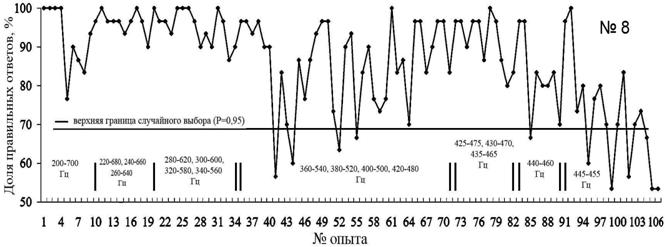
Рис. 7. Распознавание гренландским тюленем пар звуков разной частоты. (над осью обозначены применявшиеся для дискриминации частоты пар звуковых сигналов. Первые три занятия звук сопровождался голосовой командой тренера).
При предъявлении пары звуков 445 – 455 Гц животное выполняло одно из движений, стараясь получить подкрепление. Таким образом, минимальный интервал уверенного распознавания двух звуков гренландским тюленем составил 20 Гц.
5.2.3 Серый тюлень
К началу исследовательской работы самка серого тюленя №5 не имела опыта по выработке условных рефлексов на статические зрительные условные сигналы и звуки чистых тонов.
Тюлень №5 является постоянным участником цирковых представлений океанариума. В эксперименте использованы двигательные реакции "стыдно" и "ладушки", выполнявшиеся ранее при предъявлении жестового сигнала тренера. В первых опытах данной серии формировали связь: "звуковой сигнал – жест тренера – выполнение адекватного движения – подкрепление". Таких занятий было 7. Количество правильных ответов составило 100%, что, скорее всего, свидетельствует о прочности связи: жест – двигательный навык, а не о возникновении связи звук – двигательный навык.
Далее были проведены 24 занятия по предъявлению пары звуков 200 Гц и 700 Гц без подкрепления жестом. В опытах №8-31 животное получило 720 предъявлений звуковых раздражителей частотой 200 и 700 Гц. В целом процент правильных ответов на протяжении всего эксперимента был выше 50%, за исключением опытов № 12, 19, что возможно связано со слабой пищевой мотивацией тюленя во время. В дальнейшем тюлень показывал высокие результаты: 100% правильных ответов в опытах № 29-31, что позволяет говорить об уверенном дифференцировании животным звуков 200 Гц и 700 Гц по относительному признаку "выше–ниже". В опытах № 32-64 определяли величину минимального частотного интервала между двумя звуковыми сигналами, распознаваемыми серым тюленем. До конца эксперимента тюлень демонстрировал хорошую динамику и высокий уровень правильных ответов. Животное с высокой точностью дифференцировало последовательно сближаемые по частоте пары звуков. К 64 опыту серый тюлень уверенно различал звуки с интервалом в 260 Гц. Однако в декабре у животного начался период гона, и эксперимент пришлось прекратить. Тем не менее, полученные результаты свидетельствуют о том, что все исследуемые тюлени способны сравнивать и различать звуки по относительному признаку ("низкий"-"высокий"). Для распознавания очередной предъявляемой пары звуков тюленям необходимо от 1 до 5 занятий. Различение (распознавание, выбор) раздражителей по относительному признаку (выше-ниже, больше-меньше и т. п.) является важнейшим показателем их способности к обобщению (Зорина, Полетаева, 2001).
Уровень правильных ответов при выработке условных рефлексов на звуки 200 и 700 Гц при работе с молодым животным (гренландский тюлень) был значительно выше, чем у половозрелых зверей. Разницу в уровне дифференцировки звуковых раздражителей можно объяснить разным опытом работы по жестовым командам. Отсутствие большого багажа двигательно-пищевых условных рефлексов у молодых животных при осуществлении тюленем выбора – каким движением следует ответить также, вероятно, значительно повлияло на скорость обучения и уровень правильных реакций.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
