
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
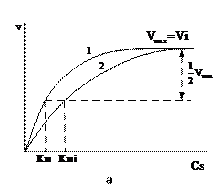
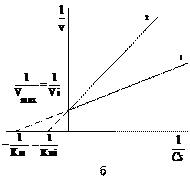
Рис. 9. Зависимость скорости ферментативной реакции от концентрации субстрата в присутствии конкурентного ингибитора (а) и ее представление в двойных обратных координатах (б). Где 1 – график без ингибитора, 2 - график с ингибитором. Vi – Vмах в присутствии ингибитора.
Неконкурентное ингибирование наблюдается, если ингибитор связывается вне активного центра. К неконкурентным ингибиторам относятся, например, тиоловые яды.
Неконкурентные ингибиторы не влияют на константу Михаэлиса, но уменьшают максимальную скорость ферментативной реакции (рис.8):
 Рис. 10. Зависимость скорости ферментативной реакции от концентрации субстрата в присутствии неконкурентного ингибитора. Обозначения как на рисунке 9.
Рис. 10. Зависимость скорости ферментативной реакции от концентрации субстрата в присутствии неконкурентного ингибитора. Обозначения как на рисунке 9.
Бесконкурентное ингибирование - ингибитор связывается только с фермент-субстратным комплексом, но не со свободным ферментом, изменяя его конформацию, что затрудняет катализ. Максимальная скорость реакции и константа Михаэлиса уменьшаются в одинаковое количество раз и на графике в двойных обратных координатах наблюдаются параллельные прямые (рис.11).
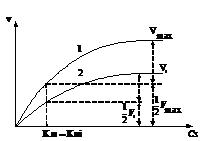
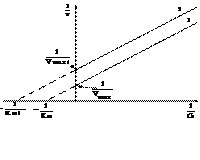 Рис. 11. Зависимость скорости ферментативной реакции от концентрации субстрата в присутствии бесконкурентного ингибитора.
Рис. 11. Зависимость скорости ферментативной реакции от концентрации субстрата в присутствии бесконкурентного ингибитора.
Смешанное ингибирование встречается, если ингибитор связывается как в активном центре, так и вне его, а комплекс ЕI сохраняет частичную активность по сравнению с нативным ферментом. Такие ингибиторы увеличивают константу Михаэлиса и уменьшают максимальную скорость ферментативной реакции. В двойных обратных координатах ситуация выглядит так (рис.12):
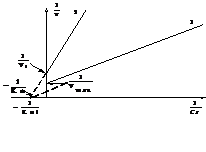
Рис.12. Представление зависимости скорости ферментативной реакции от концентрации субстрата в присутствии смешанного ингибитора в двойных обратных координатах.
Типы обратимого ингибирования ферментов представлены в таблице.
Тип ингибитора: | Механизм действия: |
КОНКУРЕНТНОЕ | I связывается в активном центре и конкурирует с субстратом. Vi max = V max ; Км i > Км |
НЕКОНКУРЕНТНОЕ | I связывается вне активного центра. Vi max < V max ; Км i = Км |
БЕСКОНКУРЕНТНОЕ | I связывается не с Е, а с комплексом ЕS. Vi max / Км i = Vmax / Км |
СМЕШАННОЕ | I связывается в активном центре и вне его. Vmax и Км уменьшаются независимо друг от друга. |
7. Классификация ферментов
Стремительное развитие энзимологии в 20 веке привело к тому, что остро встала проблема разработки единой классификации и унификации названий ферментов. В 1961 г. на V Международном биохимическом конгрессе в Москве была утверждена современная классификация ферментов, в основе которой лежит их разделение на шесть классов в зависимости от типа катализируемой реакции.
1) Оксидоредуктазы катализируют окислительно-восстановительные реакции.
Пример: исп.

В большинстве случаев дегидрогеназы катализируют обратимые реакции
2) Трансферазы катализируют реакции межмолекулярного переноса различных групп атомов,
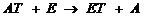
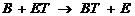
где Т - транспортируемая группа,
АТ – субстрат – донор,
В – субстрат - акцептор транспортируемой группы.
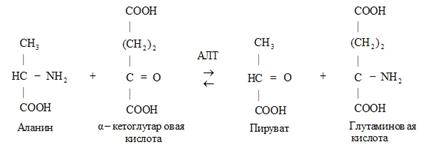
Пример 1 – аминотрансферазы, переносят альфа-аминогруппу аминокислот на место альфа-кетогруппы в кетокислотах. На схеме АЛТ – аланинаминотрансфераза.
Пример 2 - один из наиболее распространённых видов посттрансляционной модификации белка (синтез фосфопротеинов) — фосфорилирование, которое катализируют фосфотрансферазы (киназы), осуществляющие перенос фосфатной группы от молекулы аденозинтрифосфата (АТФ) на различные субстраты.
3) Гидролазы катализируют расщепление внутримолекулярных связей с присоединением воды по разорванной связи:
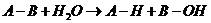 , где А-В - субстрат
, где А-В - субстрат
В качестве примеров гидролаз можно привести протеиназы, катализирующие расщепление белков и пептидов; эстеразы, гидролизующие сложноэфирные связи, гликозидазы, разрывающие гликозидные связи с присоединением воды. Все пищеварительные ферменты относятся к классу гидролаз (некоторые из них: пепсин, трипсин, химотрипсин, амилаза, липаза, рибонуклеаза).
4) Лиазы катализируют разрыв и синтез связей С-О, С-N, С-C, а также обратимые реакции негидролитического отщепления групп с образованием двойной связи.
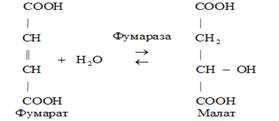
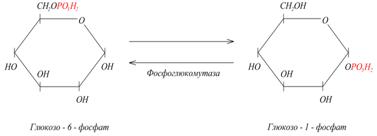
5) Изомеразы. К классу изомераз относят ферменты, катализирующие обратимые взаимопревращения изомеров. В качестве примера приведем следующую реакцию:
6) Лигазы (синтетазы) катализируют реакции синтеза различных веществ с использованием энергии АТФ или других макроэргических молекул. В качестве примера можно привести синтез карбамоилфосфата.

На основании приведенной системы классификации ферментов (КФ) был издан список ферментов, где каждому ферменту присвоен четырехзначный номер (номенклатура ферментов). Первая цифра номера указывает на принадлежность фермента к одному из шести классов. В пределах классов ферменты группируются в подклассы и подподклассы в соответствии с особенностями катализируемых реакций, четвертое число — порядковый номер фермента в его подподклассе.
Например, кислая фосфатаза имеет шифр 3.1.3.2; это означает, что она относится к классу гидролаз (3.), подклассу этих ферментов, действующих на сложноэфирные связи (3.1.), к подподклассу ферментов, гидролизующих моноэфиры фосфорной кислоты (3.1.3.), а порядковый номер фермента в данном подподклассе — 2 (3.1.3.2).
8. Внутриклеточная регуляция ферментативной активности
Регуляция ферментативной активности за счет изменения
количества молекул фермента в клетке путем биосинтеза ферментов de novo и деградации белковых молекул.
Если какой-либо белок частично разрушается и не может выполнять свои биологические функции, то с ним связывается небольшой белок убиквитин, который транспортирует дефектный белок внутрь протеасом, где убиквитин отделяется, а белок подвергается гидролизу до свободных аминокислот. Деградация белковых молекул в клетке происходит постоянно, и при замедлении синтеза фермента его внутриклеточная концентрация будет понижаться.
Регуляция синтеза ферментов осуществляется теми же путями, что и других белков. Важную роль в регуляции играют гормоны, а также условия, в которых находится организм, в частности, характер питания. Например, при переходе к пище, богатой белками возрастает скорость синтеза ферментов белкового обмена (скорость синтеза гистидазы увеличивается на 20%), а при переходе к бедной жирами диете на 30% быстрее синтезируются ферменты, входящие в комплекс синтеза высших жирных кислот.
Количество фермента в клетке зависит и от наличия продукта реакции, катализируемой данным ферментом, причем продукт может вызывать торможение синтеза фермента путем репрессии, а субстрат действует на системы, активирующие синтез. Вышесказанное можно кратко суммировать следующей схемой:
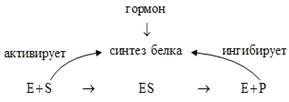
Типы регуляции ферментативной активности без изменения количества молекул фермента
Эти типы регуляции основаны на изменении активности ферментов, которые могут происходить при воздействии многочисленных факторов, которые будут рассмотрены ниже.
Влияние концентрации субстрата
При недостатке субстрата фермент не может проявлять свою активность полностью. Скорость ферментативной реакции возрастает с увеличением концентрации субстрата согласно уравнению Михаэлиса-Ментен. Рост концентрации субстрата в клетке может быть результатом интенсификации его синтеза, увеличения внеклеточных концентраций или действия регулируемых мембранных белков-переносчиков.
Иногда субстрат может выступать в роли аллостерического активатора фермента. В этом случае зависимость скорости ферментативной реакции от концентрации субстрата не подчиняется уравнению Михаэлиса-Ментен, а график этой зависимости представляет S-образную кривую. Примером аллостерической активации субстратом является влияние глюкозы на активность гексокиназы:
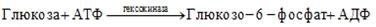
Влияние концентрации кофермента, кофакторов
Скорость ферментативной реакции прямо пропорциональна концентрации фермента. Однако, могут наблюдаться ситуации, когда ферменты не имеют возможности полностью проявлять свою активность. Например, если в организме не хватает витаминов, то не синтезируется достаточное количество коферментов, тогда даже при наличии достаточного количества молекул апофермента будет наблюдаться заниженная ферментативная активность. Такая же ситуация может иметь место при недостаточности ферментов синтеза коферментов из витаминов.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
