Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
I – распространение деполяризации в немиелинезированных волокнах;
В немиелинезированных волокнах сначала в возбужденном участке возникает возбуждение (потенциал действия, деполяризация). В этом месте проницаемость мембраны на ионы Nа+ увеличивается, поэтому его наружная часть заряжается отрицательно, а внутренняя – положительно.
А невозбужденная часть мембраны нервного волокна сохраняет свою обычную разность потенциалов: наружная среда заряжена положительно, а внутренняя – отрицательно.
Между возбужденной и невозбужденной областями возникают местные токи. Это приводит к деполяризации соседнего участка, который, в свою очередь, деполяризует следующий. Возникшая разность потенциалов смещает ионы вдоль нервного волокна внутри и снаружи от положительного заряда к отрицательному.
При этом на ранее возбужденном участке начинается стадия реполяризии. Таким образом, отрицательный заряд распространяется вдоль мембраны только в одном направлении.
В миелинизированных нервных волокнах непрерывное проведение нервного импульса невозможно. Возбуждение (деполяризация) может возникать не по всей длине мембраны, а только в перехватах Ранвье. Деполяризация одного перехвата вызывает деполяризацию соседнего.
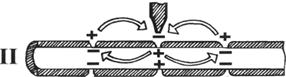 |
II – распространение деполяризации в миелинезированных волокнах;
Процессом распространения возбуждения называют движение отрицательного заряда (деполяризация) по всему нервному волокну.
При возбуждении миелинового волокна миелиновый слой не проводит электрические величины, поэтому отрицательный заряд возникает только на перехватах Ранвье.
Возникающий потенциал действия в 5–6 раз превышает порог, необходимый для возникновения возбуждения в следующем перехвате Ранвье.
Механизм распространения возбуждения по миелинизированным волокнам называется скачкообразным или сальтаторным.
Сальтаторный механизм распространения выгоднее непрерывного, так как позволяет увеличить скорость проведения нервного импульса и более экономичен с энергетической точки зрения.
Особенности сальтаторного механизма распространения:
1. Деполяризуются только небольшие участки нервного волокна.
2. Возникают меньшие потери ионов, следовательно, клетке приходится расходовать меньше энергии для обеспечения работы, Nа+,К+ – насосов.
Скорость проведения возбуждения по немиелинизированному волокну диаметром 1 мкм составляет только 2 м/с, а для волокон диаметром 0,5-1 мм эта величина уже достигает 20 м/с.
Возбуждение может распространяться не только через один, а через два перехвата Ранвье. Сальтаторное распространение импульса в миелиновых волокнах обеспечивает увеличение скорости возбуждения. При этом нервные волокна мало используют энергию нейрона.
Для миелинизированных волокон скорость проведения возбуждения зависит от длины межперехватных участков Ранвье. Возбуждение от одного перехвата к другому передается за 0,07 мс.
В то же время длина межперехватных участков Ранвье пропорциональна диаметру волокна. Поэтому, измеряя скорость проведения нервного импульса, можно оценить его функции. Если нарушается функция миелиновой оболочки, тогда скорость распространения возбуждения снижается.
При некоторых из аутоиммунных заболеваний, например «Рассеянный склероз», иммунная система организма разрушает миелиновую оболочку, т. е. происходит оголение (демиелинизация) нервных волокон. Проведение нервных импульсов через пораженный участок нарушается, что приводит к различным проявлениям: нарушению зрения и координации, мышечной слабости и др.
4. Иллюстративный материал: Презентация, слайды.
5. Литература:
1. Ф. и др. Биофизика.– М.– 2000.
2. А., Н. Биофизика.– У.– 2004.
3. Сәтбаева Х.Қ. және т. б. Адам физиологиясы –А.–2005.
4. Б. Биофизика. – Т.1,2.– М.– 1987.
5. А. и др. Биофизика.– М.– 1983.
6. И., А. Медицинская биофизика.– М.– 1978.
7. В. Биофизика.– М.– 1978.
8. А. и др. Мед. и биологическая физика.–Новгород – 2001.
6. Контрольные вопросы (обратной связи):
1. Какие виды нервных волокон Вы знаете?
2. Как образуется потенциал действия нервного волокна?
ЛЕКЦИЯ №7
1. Тема лекции: Потенциал действия кардиомиоцитов. Ионные насосы миокардиальных клеток. Распространение потенциалов действия по тканям сердца.
2. Цель лекции: обяснить студентам механизм генерации потенциала действия кардиомиоцита и проведение потенциалов действия по тканям сердца.
План лекции:
Варота |
1. Возникновение потенциала действия в кардиомиоците.
2. Ионные насосы миокардиальных клеток.
3. Характерные фазы потенциал действия клетки миокарда.
4. Основные методы изучение процессы возбуждения кардиомиоцита.
3. Тезисы лекции:
Кардиомиоцит - структурная единица сердечной мышечной ткани.
Виды кардиомиоцитов:
1) сократительные (типичные),
2) проводящие (атипичные),
3) секреторные.
1. Сократительные кардиомиоциты – это клетки цилиндрической формы, длиной до 100-150 мкм и диаметром 10-20 мкм. Они образуют основную часть миокарда, соединены друг с другом в цепочки основаниями цилиндров и обеспечивают силу сокращения сердечной мышцы.
2. Проводящие кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они генерируют электрические импульсы и передают их на сократительные кардиомиоциты, при этом обеспечивают автоматическое сокращение миокарда.
3. Секреторные кардиомиоциты находятся в правом предсердии.
Они характеризуются слабым развитием сократительного аппарата. Выполняют эндокринную функцию.
Потенциал действия мышечной клетки сердца отличается от потенциала действия нервного волокна и клетки скелетной мышцы, прежде всего длительностью возбуждения – деполяризации.
Если длительность потенциала действия аксона составляет 1 мс, клетки скелетной мышцы 2–3 мс, то длительность потенциала действия клетки сократительного миокарда желудочка и сердца составляет 250–300 мс. Это позволяет осуществить синхронное возбуждение и сокращение структур сердца для обеспечения выброса крови.
Такие особенности ПД кардиомиоцита обеспечиваются распределением ионов внутри и наружи клетки.
Распределение ионов «К», «Na» в кардиомиоцитах близко к распределению этих ионов в скелетной мышце. Однако в кардиомиоците при формировании потенциала действия и в процессе сокращения существенную роль играют и ионы кальция.
Концентрация свободных ионов кальция снаружи клетки составляет около 2 ммоль/л, но внутри клетки их концентрация очень мала: 10-4 ммоль/л.
При сокращении, концентрация свободных ионов кальция внутри клетки может возрастать до 1000 ммоль/л, но в фазе реполяризации избыток этих ионов удаляется из клетки.
Ионные насосы миокардиальных клеток. Сохранение ионного баланса в кардиомиоцитах обеспечивют К![]() – Na
– Na![]() – и Са
– и Са![]() –насосы, активно перекачивающие ионы Na
–насосы, активно перекачивающие ионы Na![]() – и Са
– и Са![]() наружу, а ионы К
наружу, а ионы К![]() – внутрь клетки.
– внутрь клетки.
Работу этих насосов обеспечивают ферменты – К![]() – Na
– Na![]() – АТФ–аза и Са
– АТФ–аза и Са![]() – АТФ–аза, находящиеся в сарколемме миокардиальных клеток.
– АТФ–аза, находящиеся в сарколемме миокардиальных клеток.
Число циклов насоса оценивается 20–ю секундами. Тогда на 1 см![]() за одну секунду происходят 2•10
за одну секунду происходят 2•10![]() циклов насосов. Так как за каждый цикл насос переносит 3 иона Na
циклов насосов. Так как за каждый цикл насос переносит 3 иона Na![]() , то всего переносится 6 •1012 ионов за 1 с на 1 см
, то всего переносится 6 •1012 ионов за 1 с на 1 см![]()
В покое проницаемость мембраны для ионов Na![]() – и Са
– и Са![]() мала, также мало, и концентрация ионов Са
мала, также мало, и концентрация ионов Са![]() вне клетки. Поэтому потенциал покоя, как и в нервных волокнах, определяется в основном разностью концентраций ионов К
вне клетки. Поэтому потенциал покоя, как и в нервных волокнах, определяется в основном разностью концентраций ионов К![]() по обе стороны клеточной мембраны.
по обе стороны клеточной мембраны.
Потенциал действия клетки миокарда имеет три характерные фазы:
І фаза – деполяризация. В этой фазе резко увеличивается проницаемость мембраны для ионов натрия. При возбуждении ее значение достигает Р=1:20. Порог активации натриевых каналов примерно достигает 60 мВ, а время жизни 1-2 мс и может доходить до 6 мс.
II фаза – плато. Эта фаза характеризуется медленным спадом потенциала действия от пикового значения ~ 30 мВ до нуля. В этой фазе одновременно работают два типа каналов - медленные кальциевые и калиевые каналы.
Кальциевые каналы имеют порог активации около 30 мВ, а время их жизни примерно 200 мс. В результате открывания кальциевых каналов возникает деполяризующий медленный входящий в клетку кальциевый ток.
Этот ток обеспечивает пассивный перенос ионов в соответствии с градиентом электрохимического потенциала.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 |
 Основные порталы (построено редакторами)
Основные порталы (построено редакторами)