

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Казахский Национальный Университет им. аль-Фараби
Факультет: химий и химической технологий
Кафедра: химии природных соединений
СРСП2
Тема: Тип биологической мембраны. Проблемы ионизаций и липофильности, биологический активные соединения.
Выполнила:
Проверила:
Алматы
2016
Содержание
1.Ведение
2. Химический состав и строение биологических мембран.
3. Липофильность и ионизация
4. Список литературы
Введение
Мембраны биологические(лат. membrana оболочка, перепонка) — функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки, а также образующие единую внутриклеточную систему канальцев, складок, замкнутых областей.
Биологические мембраны имеются во всех клетках. Их значение определяется важностью функций, которые они выполняют в процессе нормальной жизнедеятельности, а также многообразием заболеваний и патологических состояний, возникающих при различных нарушениях мембранных функций и проявляющихся практически на всех уровнях организации — от клетки и субклеточных систем до тканей, органов и организма в целом.
Мембранные структуры клетки представлены поверхностной (клеточной, или плазматической) и внутриклеточными (субклеточными) мембранами. Название внутриклеточных (субклеточных) мембран обычно зависит от названия ограничиваемых или образуемых ими структур. Так, различают митохондриальные, ядерные, лизосомные мембраны, мембраны пластинчатого комплекса аппарата Гольджи, эндоплазматического ретикулума, саркоплазматического
Химический состав и строение биологических мембран.
Состав биологических мембран зависит от их типа и функций, однако основными составляющими являются липиды и белки, а также углеводы (небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса).
Липиды. В составе биологических мембран обнаружены липиды трех классов: фосфолипиды, гликолипиды и стероиды. В мембранах животных клеток более 50% всех липидов составляют фосфолипиды — глицерофосфолипиды (фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит) и сфингофосфолипиды (производные церамида, сфингомиелин). Гликолипиды представлены цереброзидами, сульфатидами и ганглиозидами, а стероиды — в основном холестерином (около 30%). В липидных компонентах биологических мембран содержатся разнообразные жирные кислоты, однако в мембранах животных клеток преобладают пальмитиновая, олеиновая и стеариновая кислоты. Основную структурную роль в биологических мембранах играют фосфолипиды. Они обладают выраженной способностью формировать двухслойные структуры (бислои) при смешивании с водой, что обусловлено химической структурой фосфолипидов, молекулы которых состоят из гидрофильной части — «головки» (остаток фосфорной кислоты и присоединенная к нему полярная группа, например холин) и гидрофобной части — «хвоста» (как правило, две жирно-кислотные цепи). В водной среде фосфолипиды бислоя расположены таким образом, что жирно-кислотные остатки обращены внутрь бислоя и, следовательно, изолированы от окружающей среды, а гидрофильные «головки» —наоборот, наружу. Липидный бислои представляет собой динамичную структуру: образующие его липиды могут вращаться, двигаться в латеральном направлении и даже переходить из слоя в слой (флип-флоп переход). Такое строение липидного бислоя легло в основу современных представлений о структуре биологических мембран и определяет некоторые важные свойства биологических мембран, например способность служить барьером и не пропускать молекулы веществ, растворенных в воде (рис.). Нарушение структуры бислоя может привести к нарушению барьерной функции мембран.
Холестерин в составе биологических мембран играет роль модификатора бислоя, придавая ему определенную жесткость за счет увеличения плотности «упаковки» молекул фосфолипидов.
Гликолипиды несут разнообразные функции: отвечают за рецепцию некоторых биологически активных веществ, участвуют в дифференцировке ткани, определяют видовую специфичность.
Белки биологических мембран исключительно разнообразны. Молекулярная масса их в большинстве своем составляет 25 000 — 230 000.
Белки могут взаимодействовать с липидным бислоем за счет электростатических и (или) межмолекулярных сил. Они сравнительно легко могут быть удалены из мембраны. К такому типу белков относят цитохром с (молекулярная масса около 13 000), обнаруживаемый на наружной поверхности внутренней мембраны митохондрий.
Эти белки называются периферическими, или наружными. Для других белков, получивших название интегральных, или внутренних, характерно то, что одна или несколько полипептидных цепей оказываются погруженными в бислои или пересекают его, иногда не один раз (например, гликофорин, транспортные АТФ-азы, бактериородопсин). Часть белка, контактирующая с гидрофобной частью липидного бислоя, имеет спиральное строение и состоит из неполярных аминокислот, в силу чего между этими компонентами белков и липидов происходит гидрофобное взаимодействие. Полярные группы гидрофильных аминокислот непосредственно взаимодействуют с примембранными слоями, как с одной, так и с другой стороны бислоя. Молекулы белков, как и молекулы липидов, находятся в динамическом состоянии, для них также характерна вращательная, латеральная и вертикальная подвижность. Она является отражением не только их собственной структуры, но и функциональной активности. что в значительной степени определяется вязкостью липидного бислоя, которая, в свою очередь, зависит от состава липидов, относительного содержания и вида ненасыщенных жирно-кислотных цепей. Этим объясняется узкий температурный диапазон функциональной активности мембраносвязанных белков.
Число мембранных ферментов в клетке достаточно велико, однако их распределение в различных типах биологических мембран неодинаково. Некоторые ферменты (маркерные) присутствуют только в мембранах определенного типа (например, Na, К-АТФ-аза, 5-нуклеотидаза, аденилатциклаза — в плазматической мембране; цитохром Р-450, НАДФН-дегидрогеназа, цитохром в5 — в мембранах эндоплазматического ретикулума; моноаминоксидаза — в наружной мембране митохондрий, а цитохром С-оксидаза, сукцинат-дегидрогеназа — во внутренней; кислая фосфатаза — в мембране лизосом).
Углеводы в биологических мембранах находятся в соединении с белками (гликопротеины) и липидами (гликолипиды). Углеводные цепи белков представляют собой олиго - или полисахаридные структуры, в состав которых входят глюкоза, галактоза, нейраминовая кислота, фукоза и манноза. Углеводные компоненты биологических мембран открываются в основном во внеклеточную среду, образуя на поверхности клеточных мембран множество ветвистых образований, являющихся фрагментами гликолипидов или гликопротеидов. Их функции связаны с контролем за межклеточным взаимодействием, поддержанием иммунного статуса клетки, обе
Липофильность и ионизация
Перенос через мембраны лекарственных веществ с переменной ионизацией (уравнение ионизации Гендерсона-Гассельбальха). Принципы управления переносом.
Все ЛС - слабые кислоты или слабые основания, имеющие свои значения константы ионизации (рК). Если значение рН среды равно значению рК лекарства, то 50% его молекул будет находится в ионизированном и 50% в неионизированном состоянии и среда для лекарства будет нейтральной.
В кислой среде (рН меньше рК), там где имеется избыток протонов, слабая кислота будет находиться в недиссоциированной форме (R-COOH), т. е. будет связана с протоном – протонирована. Такая форма кислоты незаряжена и хорошо растворима в липидах. Если рН сместиться в щелочную сторону (т. е. рН станет больше рК), то кислота начнет диссоциировать и лишится протона, перейдя при этом в непротонированную форму, которая имеет заряд и плохо растворима в липидах.
В щелочной среде, там где имеется дефицит протонов, слабое основание будет находиться в недиссоциированной форме (R-NH2), т. е. будет непротонировано и лишено заряда. Такая форма основания хорошо растворима в липидах и быстро абсорбируется. В кислой среде имеется избыток протонов и слабое основание начнет диссоциировать, связывая при этом протоны и образуя протонированную, заряженную форму основания. Такая форма плохо растворима в липидах и слабо абсорбируется.
Следовательно, Абсорбция слабых кислот протекает преимущественно в кислой среде, а слабых оснований – в щелочной.
Особенности метаболизма слабых кислот (СК):
1) желудок: СК в кислом содержимом желудка неионизирована, а в щелочной среде тонкого кишечника она будет диссоциациировать и молекулы СК приобретут заряд. Поэтому, абсорбция слабых кислот будет наиболее интенсивной в желудке.
2) в крови среда достаточно щелочная и всосавшиеся молекулы СК перейдут в ионизированную форму. Фильтр клубочков почек пропускает как ионизированные, так и неионизированные молекулы, поэтому, несмотря на заряд молекулы, СК будут выводиться в первичную мочу
3) если моча щелочная, то кислота останется в ионизированной форме, не сможет реабсорбироваться обратно в кровоток и выделится с мочой; мочи кислая, то лекарство перейдет в неионизированную форму, которая легко реабсорбируется обратно в кровь.
Особенности метаболизма слабых оснований: противоположно СК (абсорбция лучше в кишечнике; в щелочной моче подвергаются реабсорбции)
Т. о., Чтобы ускорить выведение из организма слабой кислоты мочу необходимо ощелачивать, а чтобы ускорить выведение слабого основания ее необходимо подкислить (детоксикация по Попову).
Количественную зависимость процесса ионизации лекарства при различном рН среды позволяет получить уравнение Henderson-Hasselbach:
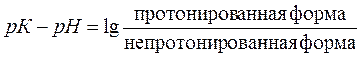 , где pKa соответствует значению pH, при котором концентрации ионизированной и неионизированной форм находятся в равновесии.
, где pKa соответствует значению pH, при котором концентрации ионизированной и неионизированной форм находятся в равновесии.
Уравнение Гендерсона-Гассельбаха позволяет оценить степень ионизации ЛС при данном значении рН и предсказать вероятность его проникновения через клеточную мембрану.
(1) Для разбавленной кислоты, A,
HA ↔ H+ + A - , где HA – концентрация неионизированной (протонированной) формы кислоты и A - - концентрация ионизированной (непротонированной) формы.
(2) Для слабого основания, B,
BH+ ↔ H+ + B, где BH+ - концентрация протонированной формы основания, B - концентрация непротонированной формы
Зная рН среды и рКа вещества, можно по вычисленному логарифму определить степень ионизации лекарства, а значит, и степень его всасывания из желудочно-кишечного тракта, реабсорбции или экскреции почками при разных значениях рН мочи и т. д.
Перенос лекарств в организме. Водная диффузия и диффузия в липидах (закон Фика). Активный транспорт.
Перенос ЛС в организме может осуществляться водной и липидной диффузией, активным транспортом, эндо - и пиноцитозом.
Особености перенос ЛС в организме водной диффузией:
1. Эпителиальные покровы (слизистые ЖКТ, полости рта и др.) – водная диффузия только очень малых молекул (метанол, ионы лития и т. д.)
2. Капилляры (кроме мозговых) – фильтрация веществ с молекулярной массой до 20-30 тыс. Да.
3. Капилляры мозга – в основном не имеют водных пор, за исключением областей гипофиза, эпифиза, зоны IV желудочка, хориоидного сплетения, медианного возвышения
4. Плацента – не имеет водных пор (хотя вопрос спорный).
5. Связывание ЛС с белками крови препятствует их выходу из кровяного русла, а значит, и водной диффузии
6. Диффузия в воде зависит от размеров молекул ЛС и водных пор
Особенности липидной диффузии:
1. Основной механизм переноса ЛС через клеточные мембраны
2. Определяется липофильностью диффундируемого вещества (т. е. коэффициентом распределения «масло/вода») и градиентом концентрации, может быть ограничена очень низкой растворимостью вещества в воде (что препятствует проникновению ЛС в водную фазу мембран)
3. Легко диффундируют неполярные соединения, трудно – ионы.
Любая диффузия (и водная, и в липидах) подчиняется закону диффузии Фика:
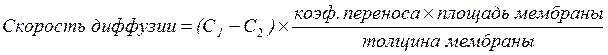 , где
, где
Скорость диффузии – количество переносимых в единицу времени молекул лекарства; С1 - концентрация вещества снаружи мембраны;С2 - концентрация вещества изнутри мембраны.
Следствие из закона Фика:
1) фильтрация ЛС тем выше, чем больше его концентрация в месте введения {S абсорбируемой поверхности в кишечнике больше, чем в желудке, поэтому абсорбция ЛС в кишечника более быстрая}
2) фильтрация ЛС тем выше, чем больше концентрация ЛС в месте введения
3) фильтрация ЛС тем выше, чем меньше толщина преодолеваемой биологической мембраны {толщина барьера в альвеолах легких значительно меньше, чем кожи, поэтому скорость абсорбции выше в легких}
Активный транспорт – перенос ЛC независимо от градиента концентрации с использованием энергии АТФ, характерен для гидрофильных полярных молекул, ряда неорганических ионов, сахаров, аминокислот, пиримидинов. Характеризуется: а) избирательностью к определенным соединениям б) возможностью конкуренции двух веществ за один транспортный механизм в) насыщаемостью при высоких концентрациях вещества г) возможностью транспорта против градиента концентрации д) затратой энергии.
Список литературы
1. Сайт http://www. biochemistry. ru/biohimija_severina/.html
2. , , и др. Гистология. М.
Медицина, 1999.
3. Сайт http://www. bibliotekar. ru/447/7.htm
6
