Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ОБЩАЯ ПАТОЛОГИЯ И ПАТОЛОГИЧЕСКАЯ ФИЗИОЛОГИЯ
Бюл. Эксп Биол и Мед 2004, 138(10): 382-385
ВЛИЯНИЕ ГИПОКСИЧЕСКОГО ПРЕКОНДИЦИОНИРОВАНИЯ КРЫС IN VIVO НА ДИНАМИКУ СОДЕРЖАНИЯ ВНУТРИКЛЕТОЧНОГО КАЛЬЦИЯ, ИНДУЦИРУЕМУЮ ДОЛГОВРЕМЕННОЙ АНОКСИЕЙ В СРЕЗАХ МОЗГА.
, ,
Институт физиологии им. РАН. Санкт-Петербург
Исследована динамика содержания внутриклеточного кальция, индуцируемая 10 мин аноксией в срезах обонятельной коры мозга крыс, предварительно подвергнутых однократному или трехкратному воздействию умеренной гипобарической гипоксии (эквивалентной подъему на высоту 5000 м). Установлено, что трехкратное прекондиционирование животных умеренной гипобарической гипоксией, в отличие от однократного, обладает нейропротектирующим эффектом. Этот эффект проявлялся в достоверном угнетении характерного патологического феномена «кальциевой перегрузки», возникающего в срезах мозга в ответ на тестирующую 10 мин аноксию in vitro.
Ключевые слова: гипобарическая гипоксия, прекондиционирование, кальций, срезы мозга.
В настоящее время интенсивно исследуется феномен «ишемической/гипоксической толерантности» мозга [2, 3, 10, 11, 17], заключающийся в том, что после умеренного гипоксического воздействия повышается резистентность нейронов мозга к последующим воздействиям более тяжелых форм гипоксии или ишемии. Методическая процедура, инициирующая гипоксическую толерантность мозга, известна как «гипоксическое прекондиционирование». Ранее нами установлено, что долговременная (10 мин) аноксия срезов обонятельной коры мозга крыс вызывает стойкую гиперактивацию NMDA рецепторов, приводящую к патологической кальциевой перегрузке нейронов [15]. Прекондиционирование срезов кратковременной аноксией, которая предъявлялась за 90 мин перед долговременной, мобилизовало срочные (также NMDA рецептор – опосредованные) механизмы толерантности и предотвращало развитие патологических процессов, индуцируемых последующей долговременной аноксией [3, 16]. В прикладном аспекте более адекватной моделью прекондиционирования представляется умеренная гипоксия in vivo. Для успешного профилактического применения гипоксического прекондиционирования in vivo важно установить его необходимую «дозу», то есть глубину и длительность прекондиционирующей гипоксии, а также период, в течение которого развивается и удерживается нейропротективный эффект, подобно тому, как это было разработано нами ранее для модели in vitro [2-4, 17].
Удобной моделью для поиска оптимальных режимов гипоксического прекондиционирования и вскрытия индуцируемых им внутриклеточных нейропротективных механизмов является двухэтапный эксперимент, в котором животное подвергают умеренной гипобарической гипоксии, а затем переживающим срезам, изготовленным из его мозга, предъявляют долговременную аноксию in vitro в качестве тестирующего воздействия [1].
В настоящем исследовании мы стремились выяснить (1) может ли прекондиционирующая умеренная гипобарическая гипоксия повысить толерантность нейронов мозга в отношении кальциевой перегрузки, вызываемой тестирующей аноксией срезов мозга экспериментальных животных, и (2) как влияет многократное гипобарическое прекондиционирование на эффективность нейропротекции по сравнению с однократным. Для решения поставленных задач была исследована динамика содержания внутриклеточного кальция, индуцированная тестирующей аноксией в срезах мозга крыс, после предъявления животным-донорам различных режимов прекондиционирующей умеренной гипобарической гипоксии.
МЕТОДИКА ИССЛЕДОВАНИЯ
Исследования проведены на крысах-самцах линии Вистар-Киото массой 190-210 г. Животные были разделены на две группы. Крыс первой группы подвергали однократному воздействию прекондиционирующей гипобарической гипоксии (1П) При этом давление в барокамере проточного типа в течение 2 часов поддерживалось на уровне 380 мм Hg, что соответствует высоте 5000 м. Во второй группе такую процедуру повторяли трижды с интервалом 24 часа (3П). Животные, не подвергавшиеся прекондиционированию, составляли контрольную группу.
Второй этап эксперимента (in vitro) проводили на следующие сутки после процедуры прекондиционирования. Животных обеих групп декапитировали и изготовляли переживающие тангенциальные срезы обонятельной коры толщиной 400 мкм с помощью вибротома (EMS-4000, Electron Microscopy Sciences). Срезы помещали в проточную инкубационную среду с температурой 37.5оС. Спектрофлуориметрическое определение содержания внутриклеточного кальция производили в инкубационных системах, приспособленных для определения динамики содержания как связанного (Са-с), так и свободного (Са2+цит) внутриклеточного кальция, для чего в различных сериях экспериментов в качестве измерительных приборов использовали контактный флуоресцентный микроскоп ЛЮМАМ КФ (ЛОМО, СПб) и спектрофлуориметор Hitachi F-2000 (Hitachi, Tokyo), соответственно. Для получения флуоресцентного сигнала, характеризующего содержание Са-с, применяли хлортетрациклиновый флуоресцентный зонд (Sigma). При этом измерялся квантовый выход флуоресценции в спектральной области с максимумом 520 нм, возбуждаемой облучением с длиной волны 410 нм. Для регистрации относительных изменений уровня Са2+цит был применен флуорохром fura-2AM (Molecular Probes, USA). Расчетной характеристикой этих изменений служило отношение флуоресцентных сигналов с максимумом 510 нм, полученных при возбуждающем облучении с длинами волн 340 нм и 380 нм, соответственно. Более детально обе флуоресцентные методики описаны ранее [15]. Аноксия in vitro создавалась путем замены кислородсодержащей инкубационной среды на азотсодержащую с соответствующей сменой газовых потоков (O2 на N2) над поверхностью инкубационных растворов в камерах. После 10 мин аноксической суперфузии нормоксические условия восстанавливались обратной манипуляцией. В обеих сериях экспериментов измерения производили каждые 5 мин в течение аноксии и 70 мин реоксигенации. Результаты представлены на графиках в виде серии средних значений ± стандартная ошибка среднего. Статистическая обработка данных проведена путем вариантного анализа (ANOVA) с применением Dunnett-теста.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
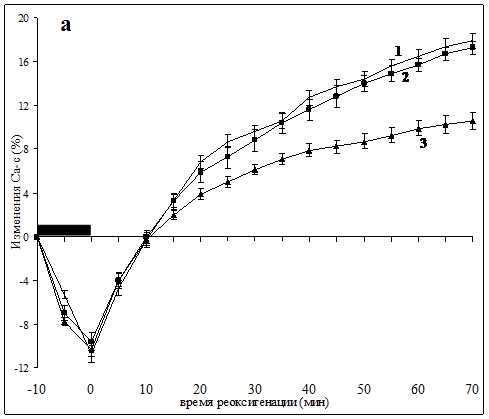
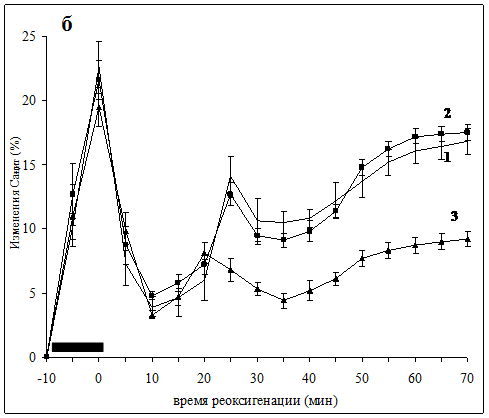
В срезах контрольных животных во время аноксии уровень Са-с снижался в среднем до 89% исходной величины, но возрастал в постаноксическом периоде, достигая 117.9±1.1% к 70 мин реоксигенации. Такая динамика содержания Са-с отражает начальный процесс высвобождения кальция из внутриклеточных секвестров в аноксической фазе, который сопровождается входом внеклеточного кальция и его частичным связыванием в фазе реоксигенации [3, 15]. В срезах, полученных от животных группы 1П, реакция не отличалась от контрольной. В то же время, в группе 3П характерное для постаноксического периода увеличение уровня Са-с было достоверно подавлено после 20 мин реоксигенации. Уровень Са-с в этой группе возрастал к 70 мин лишь до 110±0.8% (Рис. 1а). При определении динамики Са2+цит в контрольных срезах выявлены аноксическая и постаноксическая фазы повышения его уровня, непосредственно демонстрирующие развитие феномена «кальциевой перегрузки» при реоксигенации. Как и в предыдущей серии динамика содержания Са2+цит в группе 1П не отличалась от контрольной, т. е. однократное гипоксическое прекондиционирование оказалось недостаточно эффективным для индукции нейропротективных механизмов. Вместе с тем, в группе 3П постаноксическая фаза возрастания уровня Са2+цит после 20 мин реоксигенации была значительно подавлена, составив к 70 мин лишь 9.2±0.6% прироста уровня Са2+цит против 16.8±1.0% в контрольных срезах (Рис. 1б).
Таким образом, полученные результаты показывают, что доза трехкратного гипоксического прекондиционирования более эффективна, чем однократного. Возможно, увеличение числа прекондиционирующих сессий, до какого-то предела, приведет к еще большей степени нейропротекции. Поиск этого предела для данной модели является предметом дальнейших исследований.
Использованная модель прекондиционирования in vivo, при которой интервал между первой гипобарической сессией и тестирующей аноксией in vitro составляет 3 суток, очевидно, индуцирует долгосрочные механизмы толерантности нейронов. Согласно современным представлениям, такие механизмы сопряжены с экспрессией генетического аппарата клетки и усилением синтеза регуляторных белков и пептидов [2, 3, 8]. Известно, что в механизм формирования долгосрочной гипоксической толерантности нейронов вовлекаются ранние гены [5, 6, 14], белки теплового шока [7, 11], пептиды семейства нейротрофинов [13], кортикотропин-рилизинг фактор [9], опиоидные пептиды [12]. Представленные данные позволяют предположить, что нейропротективный эффект механизмов долгосрочной индуцированной толерантности нейронов к гипоксии, при условии прекондиционирующего воздействия достаточной силы, приводят к единому нейропротективному эффекту, а именно ограничению характерной для тяжелых форм гипоксии глутаматной (прежде всего, NMDA рецептор – опосредованной) нейротоксичности, проявляющейся в кальциевой перегрузке и гибели нейронов.
ЛИТЕРАТУРА
1. , , // Бюлл. Экспер. Биол. и Медицины 2001. Т.132. N6. С.1154-1156.
2. Самойлов и адаптация: молекулярно-клеточные механизмы. СПб, 1999.
3. , , и др. // Рос. Физиол. Журн им. . 2001. Т.87. N6. С.714-729.
4. , , // Рос. Физиол. Журн. им. . 1999. Т.85. N1. С.139-148.
5. Abe H., Nowak T. S.Jr. // Acta Neurobiol. Exp. 1996. Vol.56, N1. P.3-8.
6. Baron F., White R. F, Spera P et. al. // Stroke 1998. Vol.29. N12. P.1937-1951
7. Chen J., Graham S., Zhu R. et al // J. Cereb. Blood Flow Metab. 1996 Vol.16. P.566-577
8. Chen J., Simon R. P // Neurology. 1997. Vol.47. P306-311
9. Fox M. W., Anderson R. E., Meyer F. B. // Stroke. 1993. Vol.24. N7. P.1072-1075.
10. Kato H., Liu Y., Araki T., Kogure K. //. Brain Res. 1991. Vol.553. N2. P.238-242.
11. Kitagawa K., Matsumoto M., Tgaya M. et al. // Brain Res. 1990. Vol.528. N1. P.21-24.
12. Mayfield K. P., D'Alecy L. G. // Brain Res. 1992. Vol. 582, N 2. P. 226-231.
13. Pechan P. A., Yoshida T., Panahian N. et al. // Neuroreport. 1995. Vol.6. N4. P.669-672.
14. Rybnikova E., Tulkova T., Pelto-Huikko M., Samoilov M. Neurosci. Lett. 2002. Vol.329. N1. P.49-52.
15. Semenov D., Samoilov M., Zielonka P., Lazarewicz J. Resuscitation 2000. Vol.44. P.207-214.
16. Semenov D., Samoilov M., Lazarewicz J. Neurosignals 2002. Vol.11. P.329-335.
17. Vanucci R. C., Towfighi J., Vanucci S. J. J. Neurochem. 1998. Vol.71. P.1215-1220.
ПОДПИСИ К РИСУНКУ
Изменения уровня Са-с (а) и Са2+цит (б), вызванные 10 мин аноксией в срезах контрольных животных (1) и животных, подвергнутых однократному (2) или трехкратному (3) гипоксическому прекондиционированию.
Период аноксии отмечен черным прямоугольником. Для каждого графика n=7-9.