

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК. 595.782 (471.327)
МАТЕРИАЛЫ К РАСПРОСТРАНЕНИЮ, ЭКОЛОГИИ И БИОЛОГИИ ПАРУСНИКА МНЕМОЗИНЫ DRIOPA MNEMOSYNE (LINNAEUS, 1758) (LEPIDOPTERA: PAPILIONIDAE) НА ТЕРРИТОРИИ ПЕНЗЕНСКОЙ ОБЛАСТИ
О. А. ПОЛУМОРДВИНОВ, С. В. ШИБАЕВ
КАФЕДРА ЗООЛОГИИ И ЭКОЛОГИИ
ПЕНЗЕНСКОЕ ОТДЕЛЕНИЕ РУССКОГО ЭНТОМОЛОГИЧЕСКОГО ОБЩЕСТВА
Данная работа продолжает серию публикаций пензенских энтомологов (Полумордвинов, Монахов, 2003 и 2005; Полумордвинов, Шибаев, 2006), посвященных более детальному изучению биологии, экологии и распространению на территории области видов насекомых, занесенных в Красную книгу Российской Федерации (Животные, 2001) и, соответственно, Пензенской обл. (Животные, 2005). В предлагаемой работе предпринята попытка на примере территории Пензенской области, более полно охарактеризовать современное состояние популяций мнемозины - Driopa mnemosyne (Linnaeus, 1758) в Среднем Поволжье.
К сожалению, в настоящее время сложилась такая ситуация, что даже среди энтомологов-полевиков нет единого мнения не только в необходимости самих Красных книг, но и в целесообразности охраны насекомых, попавших на их страницы. Современное состояние одного из таких «банальных» - как они считают видов, мы и предлагаем рассмотреть в данной работе.
Описание бабочки. Длина переднего крыла 24-31 мм. Крылья белые, слабо опыленные. Кайма передних крыльев черная или серая (прозрачная), имеет черное дискальное пятно и недалеко от него в ячейке второе пятно. На заднем крыле черное затемнение развито по его заднему краю и доходит до ячейки [1]. Самки несколько темнее самцов. Их тело голое, лоснящееся, с желто-зелеными пятнами на боках. Тело самцов в мягком светлом опушении. Половой диморфизм выражен слабо [2]. Хочется отметить что, бабочки собранные даже в одной популяции могут быть в значительной мере изменчивы – что является индивидуальными вариантами. Рациональная внутривидовая систематика этого политипического вида может быть, вероятно, разработана на основе феногеографического анализа [3].
Замечания по систематике. Следует констатировать, что в настоящее время остается очень запутанным и неразрешенным вопрос о систематическом положении популяций мнемозины - Driopa (Parnassius) mnemosyne (Linnaeus, 1758), обитающих в европейской части России. Только на территории Восточной Европы было открыто более двух десятков подвидов (ssp.) и форм [4, 5]. По мнению этих авторов только в Поволжье было отмечено три таксона. Так из Саратова был приведен (ssp.) – P. m. craspedontis Fruhstorfer, 1909. В Пензе описан (ssp.) – P. m. weidingeri Bryk, Eisner, 1914 («Представляющего собой переход от (ssp.) craspedontis Fruhst. к (ssp.) uralca Bryk, 1921, но с более слабым чешуйчатым напылением. Полулунное пятно стекловидной полосы переднего крыла оттянуто назад. Оба пятна центральной ячейки всегда мощнее. – Самец с удлиненной, не так резко утончающейся, стекловидной полосой. Медиальное пятно заднего крыла то исчезающее, то явственное, однако никогда не такое мощное как у самцов (ssp.) uralca Bryk. – Самка с удлиненной стекловидной полосой, которая не полностью достигает заднего края крыла. Пятно заднего края переднего крыла скромное. Желтая центровка медиальных глазков на исподе крыльев обычна. Длина переднего крыла: ♂♀ - 32 мм.») [4]. А в Елабуге обитает (ssp.) – P. m. ugrumovi Bryk, 1914. Интересно что именно из этих таксонов, различными современными исследователями [6, 7, 8 и др.] наиболее часто выделяется какой то один (совершенно произвольно и неубедительно), который по их мнению населяет территорию центральной части восточно-европейской России. По мнению А. Г. Татаринова такая же запутанная ситуация с популяциями (ssp.) мнемозины сложилась и на Урале, где требуются дополнительные обоснования для обособления различных географических рас - P. m. timanica Eisner, Sedykh 1964, P. m. ugrumovi Bryk, 1914 и P. m. uralca Bryk, 1921 [2].
В данной работе мы не будем рассматривать целесообразность выделения этих таксонов (ssp.) – это дело отдельных специальных исследований. Но, исходя из своего опыта полевых исследований различных популяций мнемозины, мы вынуждены согласится со словами (Некрутенко, 1990) – «P. mnemosyne характеризуется чрезвычайно сильной индивидуальной изменчивостью. Наиболее изменчивыми признаками являются размеры и форма темных элементов крылового рисунка и полупрозрачных участков на крыльях, а также общая пигментация: окраска фона может изменяться от чисто белой до серой, дымчатой; изменчивы и размеры бабочек. Тенденции индивидуальной изменчивости имеют хорошо выраженную экологическую обусловленность. Обитатели прохладных, влажных горных биотопов отличаются более интенсивной общей пигментацией, в жарких, сухих местностях окраска всегда светлее. Анализ доступного материала (автор анализировал популяционную изменчивость мнемозины собранной в различных точках Кавказа - прим. авторов*) позволило установить, что признаки, использованные авторами при установлении … таксонов, которым первоначально был предан статус подвидов (количество описанных инфраподвидовых форм необозримо), не характеризует локальных популяций. Почти все описанные фенотипы можно обнаружить в любой популяции при одномоментном или проводимом в течение нескольких сезонов взятии адекватных выборок. Ревизия типов и обзор сведений, содержащихся в оригинальных описаниях (материалов собранных на Кавказе*), показали, что в большинстве случаев подвиды устанавливались по предельно малым выборкам (1-2 экз.) или по «нетипичным» экземплярам более крупной серии; только эти уклоняющиеся экземпляры и обозначались в качестве типов устанавливаемого подвида» [3]. Все вышесказанное можно в полной мере отнести как к нашим сборам - D. mnemosyne (L.) в Пензенской области, так и к известным нам материалам с территории Среднего Поволжья.
Распространение. Локально и спорадически: Пиренеи, горный массив Альп, от Италии и Северной Сицилии, до Юга Польши, Балканы, от Греции до Северной Турции. Северо-Восточ. Европа и Балтика [9]. Европейская часть России, Передняя и Ср. Азия, Вост. Казахстан, Ср. и Юж. Урал, Зап. Сибирь (Тюменская обл.) [8].
Пензенская обл.: На территории области известно обитание вида в 18 районах [10, 11, 12]. Названия растений приведены по монографии А. А. Солянова [13]. Использованы общепринятые сокращения и [*] для дополнительных комментариев коллекторов (+ важных). Для более точной привязки к местности точек сбора полевого материала, использовался наиболее информативный (есть список населенных пунктов) Атлас Пензенской области (М. 1:200 000) [14].
МАТЕРИАЛ: г. Пенза: 1 ♂, «г. Пенза, гор. лес», 17.VI.1968 (Левкович); 1 ♀, 1) «Согласье» [лес на северо-востоке г. Пензы*], 17.VI.1969 (Левкович); «С/х техн., 13.VI.1965 (О. В. Шлыков) [15]; 1 экз., «окр. г. Пензы», 1970 (Ю. Н. Стариков); 2) 2 ♀ (видел еще несколько), западная опушка и поляны «Арбековского леса», 20.VI.1998 (А. Н. Роганин), [ + ряд наблюдений в V-VI.1999 и 2000*]; + довольно большие сборы различными коллекторами в период с 1960 по 1990 гг.
Многочисленные стационарные наблюдения за популяциями проводились: 3) в 0,5 - 2 км. южнее д/п. «Барковка», опушки и поляны дубового леса в пойме р. Суры (правый, восточный берег), 20.V. – 22.VI. с 1999 по 2006 (О. А. Полумордвинов, А. Н. Роганин, Д. Н. Подгорнов и др.); 4) «Ахуны»: пойменный лес р. Суры, от 2 км. севернее и до 2 км. юго-восточнее, лиственный лес д/п. «Бурчиха», 10.V – 15.VI. с 2000 по 2006 (С. В. Шибаев); 5) смешанный лес в окр. г. Заречного (восточнее г. Пензы), V-VI. 2000 - 2005 (С. В. Шибаев).
Пензенский р-он.: 6) ♂♂ и ♀♀ (в полдень, при ярком солнце наблюдали массовый выход бабочек), в 3 км. северо-западнее п. Мичуринский, опушка смешанного лиственного леса (с преобладанием дуба) и травянистого склона балки, 3.VI. 2000 (О. А. Полумордвинов, Д. В. Поликанин) [интересно что часть самок вышедших ранее, уже имели сфрагис!]; + различные сборы материала в этом же лесном массиве, V-VI. 2001-2005 (В. И. Кузнецов и Д. Н. Подгорнов); 7) ♂♂ и ♀♀, окр. с. Алферьевка, лиственные леса и дубняки, IV, V,VI. 2001-2003 (И. В. Евсикова) [в данной точке производились систематические наблюдения за экологией популяций хохлатки плотной (кормового растения гусениц) и мнемозины].
Башмаковский р-он.: 8) ♂♂ и ♀♀, от с. Знаменское (правый, коренной берег р. Буртас), по юго-западной опушке лиственного леса (с богатым разнотравьем), до д. Самариха, VI. 2002 и 2003 (И. М. Горячкина).
Бековский р-он.: 9) 1 ♂ (наблюдал большое количество экз.), южнее п. Беково и п. Сахзавод, лесные поляны и опушки дубрав в пойменном лесу р. Хопер, весь VI.1998 (А. В. Полумордвинов) [особенно часто бабочки встречались в окр. озера «Змеиного»].
Белинский р-он.: 10) 7 экз., западнее с. Чернышево, пойма р. Ворона, на опушке дубравы, 23-28.V. 2002 (ст. заоч.); 1 ♂, 1 км. северо-западнее п. Поим, опушка лиственного леса, 27.V. 2002 (А. Е. Барышев).
Бессоновский р-он.: 11) 3-7 экз., в 1-1,5 км. северо-восточнее с. Победа, опушка и поляны в смешанном лесу «Светлополянского лесничества» [по дороге «Тени Исчезают в Полдень» и правее ручьев], к. V - с. VI. 1999-2006 (О. А. Полумордвинов, А. Н. Роганин, Д. В. Поликанин и др.); 12) 1♀ (наблюдал около 30 облетанных экз.), в 2 км. северо-западнее с. Пазелки, опушка (южная) смешанного леса и на травянистых склонах у ручья, 20.VI. 2002 (С. В. Шибаев) [в поисках цветущих растений бабочки залетали на дачные участки - расположенные рядом с лесной опушкой].
Городищенский р-он.: 13) единичные экз. ♂♂ и ♀♀ (+ редкие визуальные наблюдения), о/п. «Никоново» - от 1 до 2,5 км. южнее и юго-западнее, по остаткам дубового леса на первой террасе поймы р. Сура (правый берег), 1-11.VI. 2002-2005 (С. В. Шибаев, О. А. Полумордвинов) [популяция с очень низкой численностью, бабочки окрашены контрастно].
Земетчинский р-он.: 14) окр. с. Пашково, смешанный лес, V - VI. 1989-1994 (Ю. И. Байков) [визуальные наблюдения в разные годы]; + ряд других энтомологических сборов.
Каменский р-он.: 15) 1♀ [мелкая, рисунок крыла контрастный, правое заднее кр. значительно повреждено по анальному краю в средней части] и ♂♂ (выпустил), в 1 км. восточнее д. Новая-Есинеевка, южная опушка лиственного леса (с преобладанием дуба – на месте вырубленной дубравы) на левом коренном берегу р. Варежка, 1.VI. 2002 (О. А. Полумордвинов) [популяция с очень низкой численностью за все годы наблюдений 1991-2005. Хотя популяции кормового растений гусениц изучаемого вида - хохлатки плотной, обильны]; 16) ♂♂ и ♀♀, северо-восточная окр. г. Каменки, опушка и поляны нагорной дубравы (правый коренной берег р. Атмис, район «Аэродрома»), к. V. с. VI. 1995-2004 (В. М. Великоиваненко) [именно в данной популяции были произведены основные наблюдения и видеосъемка биологии гусениц мнемозины]; + ряд других находок.
Камешкирский р-он.: 17) 2 ♂ и 1♀, в 2 км. севернее с. Бегуч, опушка смешанного леса, 11.VI. 2004 (В. Н. Тремасов); + ряд других сборов.
Колышлейский р-он.: 18) ♂♂ и ♀♀, в 1 км. юго-западнее д. Алферьевка, пойма р. Хопер (левый берег), к. V. 2002 (А. В. Леонтьев); 1 ♂ и 1 ♀, в 2 км. северо-западнее с. Трескино, поляна в широколиственном лесу, н. VI. 2002 (А. В. Курдюкова); 19) ♂♂ и ♀♀ (видел 10 экз.), в 2 км. юго-восточнее ст. Скрябино, южная опушка смешанного леса, 10.VI. 2003 (С. В. Шибаев).
Кузнецкий р-он.: 20) 1 ♀, «окр. с. Ульяновка, 15.VI. 1978» (А. В. Каретников); + ряд энтомологических сборов с неточными этикетками.
Лунинский р-он.: 21) ♂♂ и ♀♀, в 3 км. юго-западнее ст. Гольцовка, опушка и поляны (южной экспозиции) в смешанном лесу на левом коренном берегу р. Шукша, к. V - с. VI. 1999-2004 (Е. М. Монахов, А. М. Монахов) [популяция с крайне низкой численностью за все годы наблюдений]; 22) ♂♂ и ♀♀, окр. с. Иванырс, остатки дубрав в пойме р. Сура (правый берег), к. V- с. VI. 1999-2004 (Е. М. Монахов, М. А. Монахов и А. М. Монахов); 23) ♂♂ и ♀♀, окр. д. Луговое, остатки дубрав на правом коренном берегу р. Сура, VI. 2000 и 2001 (О. А. Полумордвинов, А. Е. Барышев) [16]; 24) ♂♂ и ♀♀ (наблюдал около 15 экз.), в 5 км. юго-восточнее с. Старая Кутля (окр. д. Белый Родник), смешанный лес на первой пойменной террасе р. Сура (правый берег), 2.VI. 2002 (С. В. Шибаев); + ряд других сборов.
Неверкинский р-он.: 25) ♂♂ и ♀♀ (визуальные наблюдения), севернее с. Бикмурзино, нагорный злаковый (остепненный) разреженный дубняк, правый (коренной) берег р. Кадада, территория ООПТ «Шуро-Сиран», 2.VI. 1999 (Т. Г. Стойко) [численность бабочек в популяции высокая (но разреженная), южная окраина леса со следами чрезмерного выпаса скота].
Пачелмский р-он.: 26) ♂♂ и ♀♀, по юго-восточной опушке дубняка [рубленая дубрава с примесью различных лиственных пород] от д. Новый Валовай до д. Ст. Валовай, с. и к. VI. 2004 и 2005 (Е. В. Заикина); + ряд других энтомологических сборов.
Сердобский р-он.: 27) ♂♂ и ♀♀ (наблюдали более 10 экз.), восточнее г. Сердобска по южной опушке нагорной дубравы (правый коренной берег р. Сердоба), 2.VI. 2006 (С. В. Шибаев, Е. М. Монахов, Д. Н. Подгорнов) [численность популяции мала из за систематических сборов местным населением хохлаток для букетов* личн. ком. И. В. Муравьева].
Сосновоборский р-он.: 28) ♂♂ и ♀♀, окр. п. Сосновоборск, опушки и поляны южной экспозиции в смешанном лесу, н. и с. VI. 2004-2006 (В. А. Чернышову и Д. А. Капустину); + ряд других энтомологических сборов.
Тамалинский р-он.: 29) ♂♂ и ♀♀, окр. с. Зубрилово (пойма р. Хопер), к. V - с. VI. 1960-70 гг. (Ю. Н. Стариков); + ряд других энтомологических сборов.
Шемышейский р-он.: 30) ♂♂ и ♀♀, окр. «Биостанции ПГПУ» («Кордон»), в 9 км. севернее п. Шемышейка, поляны в смешанном лесу (преобладают лиственные породы), к. V - с. VI. 1991 – 2005 (Т. Г. Стойко, Н. Ф. Золина) [многочисленные наблюдения и редкие сборы бабочек студентами на полевой практике по зоологии беспозвоночных животных].
+ Достаточно многочисленные энтомологические сборы бабочек мнемозины с неточными или очень краткими этикетками.
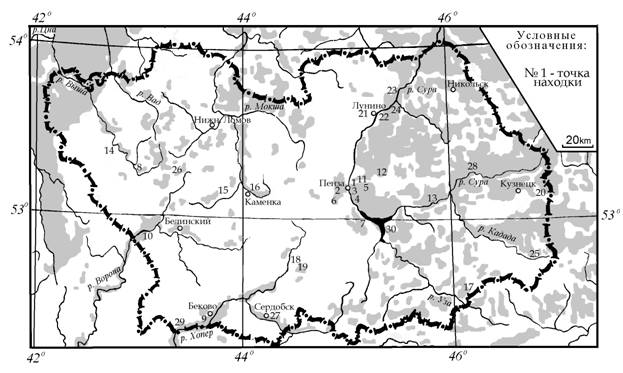
Карта № 1. Карта-схема распространения и мест находок (1960-2006 гг.) мнемозины - D. mnemosyne (L.), на территории Пензенской области.
Очевидно, что в настоящее время вид обитает во многих районах Пензенской области. На основании приведенного кадастра распространения вида и ряда энтомологических материалов (1960 – 2006) не вошедших в него, была составлена карта-схема (Карта № 1) современного распространения - Driopa (Parnassius) mnemosyne (Linnaeus, 1758) на территории Пензенской области [12].
Места обитания, особенности биологии и экология. В области вид заселяет широколиственные и производные от них смешанные леса. Обитание локальных популяций в основном отмечено на южных и юго-восточных опушках нагорных дубрав и производных от них дубняков (с порослевым возобновлением): на водоразделах, по коренным берегам мелких рек и по пойменным террасам крупных рек области – Сура, Хопер и Ворона. Интересно, что энтомологом Л. В. Большаковым в Тульской области популяции мнемозины отмечены в основном для полосы хвойно-широколиственных лесов долины р. Оки [17]. Тогда как в Пензенской (наши данные), вид практически исчезает в лесах с преобладанием сосны, что объясняет его достаточную редкость в Засурье. Это согласуется и с данными ботаников (Заугольновой, Морозовой, 2004) - «Известно, что при хорошей сохранности широколиственных лесов высоким постоянством обладают виды весенних эфемероидов (…Corydalis solida,…), которые слабо представлены в хвойно-широколиственных и широколиственно-хвойных лесах» [18]. В области бабочки встречаются в основном на хорошо прогреваемых лесных опушках, полянах, просеках, дорогах и прилегающих к лесам луговых склонах. Как правило в биотопах, где произрастает единственное известное у нас кормовое растение гусениц исследуемого вида – хохлатки плотной (Corydalis solida (L.) Clairv) [12]. Несмотря на специально предпринятые нами поиски, бабочки остались неизвестными в местах произрастания других видов хохлаток – х. маршала (C. marschalliana (Pall.) Pers.) и тем более маловероятны на х. полой (C. cava (L.) Schweigg. еt Koerte.) [13].
Вызывает искреннее удивление что многие исследователи булавоусых чешуекрылых (Lepidoptera: Rhopalocera) не имея собственных наблюдений переписывают друг у друга неподтвержденные сведения о том, что гусеница Driopa (Parnassius) mnemosyne (L.) – питается ночью (!?). За время наших полевых исследований (1995-2006) питание гусениц было отмечено – днем (!) в теплую погоду (чаще при ярком солнечном освещении) [12]. Это нашло подтверждение и в работе А. Г. Татаринова (1999) для популяций мнемозины, которые он изучал на европейском Северо-Востоке России [2]. Интересно, что в случае беспокойства (опасности): шум сухой листвы под ногами, резкие звуки, или даже при резком изменение освещения (напр. от тени наблюдателя) – гусеница стремится быстро скрыться в сухом лиственном опаде. Вероятно, эта индивидуальная особенность выработалась в результате уязвимости гусеницы в весеннем широколиственном лесу (поздно одевающемся в листву) на открытом, светлом и хорошо просматриваемом участке, где произрастает их эфемероидное кормовое растение. По данным (Смирновой, Чарушиной, 1975) C. solida отличается светолюбием и для нормальной продуктивности фотосинтеза, необходима освещенность более 50 % от полной [19]. Очевидно, что питание гусеницы в дневное время обусловлено еще и тем, что для территории области в апреле-мае обычна не только прохладная погода, но и не редки в ночное время почвенные заморозки (вероятен и снег). А показатель положительных среднесуточных температур апреля и начала мая в ночное время составляет всего: + 1º, + 3º (среднестатистическая tº) [20]. А если еще учесть, что зона активной жизни насекомого лежит в пределах от + 3 до + 40º (Ушатинская, 1957), то нормальный обмен веществ (при физиологических процессах) у гусеницы маловероятен [21]. Если сопоставить наши полевые исследования и данные многолетних фенологических наблюдений М. И. Чинаева и Л. В. Цинговатова (1928-1960) [22], то можно сказать что: а) вегетация хохлатки плотной начинается при переходе среднесуточной температуры воздуха + 5 º (30.III. – 20.IV); б) зацветание, при установлении tº + 10 º (18.IV – 4.V); в) зеленение и зацветание дуба чересчатого - Quercus robur L. (30.IV – 17.V); г) последние заморозки на почве приходятся на цветение черемухи обыкновенной (Padus avium Mill.) и могут иметь место до 17.V – 1.VI (!); д) выход первых бабочек - D. mnemosyne отмечен при установлении среднесуточной tº + 15 º (в среднем с 30.V). Таким образом, становится совершенно очевидно, что нормальное развитие гусеницы исследуемого вида в климатических условиях Пензенской области (следовательно и Среднего Поволжья) практически невозможно в ночное время. В дополнение хочется отметить тот факт, что окраска гусеницы исследуемого вида – черная или темно-серая, с обоих боков идет по одному ряду оранжево-красных пятен. То есть темная окраска напрямую способствует повышению температуры ее тела, за счет дополнительного нагрева от солнечных лучей. Развитие гусеницы (20-25 дней) завершается в середине - конце мая, окукливание в плотном коконе на почве, или в травяном опаде. Стадия куколки продолжается примерно две недели [1, 2, 3, 8, 9, 12].
Лет бабочек в одном поколении, с середины мая до конца июня [12]. Интересно, что из за различия среднесуточных температур воздуха и разной степени инсоляции земной поверхности (следовательно и вегетации растений) [20, 22], выход бабочек в лесостепи юго-запада Пензенской обл. начинается на неделю раньше, чем в лесах северо-востока. Бабочки активны в ясную солнечную погоду (в основном с 11 до 16 часов), полет медленный, планирующий. Утром или в пасмурную погоду плохо и неохотно летают, напуганные (отмечено в основном утром и у недавно отродившихся бабочек) падают в траву и лежа на почве, начинают издавать отпугивающее «пощелкивание» (за счет трения лапок и нижней поверхности крыльев). В результате многочисленных наблюдений для области выявлены следующие кормовые растения [13] бабочек (имаго). В скобках указаны точки находок (Карта 1):
Смолевка клейкая – Silene viscosa (L.) Pers. (3) Смолка клейкая – Steris viscaria (L.) Rafin. (4) Фиалка удивительная – Viola mirabilis L. (4) Земляника зеленая – Fragaria viridis Duch. (3, 4) Черемуха обыкновенная – Padus avium Mill. (4) Горошек заборный – Vicia sepium L. (24) Чина гороховидная – Lathyrus pisiformis L. (24) Вероника дубравная – Veronica chamaedris L. (3, 4) Марьянник дубравный – Melampyrum nemorosum L. (4) Подорожник средний – Plantago media L. (4) Черноголовка обыкновенная – Prunella vulgaris L. (4) Будра плющевидная – Glechoma hederacea L. (3) Одуванчик лекарственный – Taraxacum officinale Wigg. (3, 4) Нивяник обыкновенный – Leucanthemum vulgare Lam. (4)Большая часть из перечисленных выше растений многолетники. Часть из них медоносы. В результате длительных наблюдений выявлено хорошее зрение у бабочек, выражавшееся в быстрой реакции на цветки и соцветия кормовых растений, замеченных ею в полете. Какого либо предпочтения кормится цветущим растениям с определенным спектром цветка, не выявлено. Бытующее мнение, о несъедобности (ядовитости?) взрослых бабочек, в лабораторных условиях не подтвердилась. Спаривание отмечено в первые дни после выхода бабочек из куколок, после чего у самок на нижней стороне вершины брюшка образуется особый ногтевидный белый придаток – сфрагис. Остается не ясным способность бабочек (самок) к точному поиску мест, где произрастает весеннезеленое кормового растение гусениц (!?). По данным различных наблюдателей откладывание яиц самкой середина-конец июня, происходило на почву, совершенно точно в тех местах, где в апреле и начале мая произрастала - Corydalis solida (L.) Clairv (!?). От самих растений (вегетативной части) в это время практически уже не остается и следа. Факты рассеивания яиц неизвестны и крайне сомнительны. Зимует яйцо со сформировавшейся гусеницей [2, 8]. Мечение имаго (бабочек) в полевых условиях цветными (водостойкими) маркерами и наблюдение за ними показали, достаточную консервативность особей выразившуюся в относительной локальности (особенно самцы) их мест обитания 1 – 1,5 км². При этом у бабочек данного вида обнаружилась интересная особенность – они не склонны перелетать (вылетать) через (на) открытые участки местности: поля, большие луга, пустоши, крупные вырубки, овраги и балки.
Численность и лимитирующие факторы. С XVIII века наблюдается прогрессирующее уменьшение площади ареала и неуклонное сокращение общей численности вида, как в Европе, так и в России [1, 2, ]. Интересно, что основной ареал – D. mnemosyne (L.) практически совпадает с распространением основного кормового растения гусениц – хохлатки плотной C. solida (L.) Clairv [23]. Показательно, что в начале ХХ века вид уже не указывался как «наичаще встречающийся в Пензенской губернии» (Попов, 1901), тогда как другие четыре вида наших парусников (P. machaon, I. podalirius, Z. polyxena, P. apollo) в нем приводились [24]. В настоящее время для территории Пензенской обл. намечается неуклонная тенденция к сокращению и фрагментации ареала мнемозины, особенно в западных и юго-западных аграрных районах области [12]. Если проанализировать карту-схему (Карта № 1) распространения вида в области, очевидным становится факт, что большинство популяций D. mnemosyne у нас сохранилось в виде локалитетов, обитающих в водоохранных зонах рек. Снижению численности вида и его локализации, способствуют следующие антропогенные факторы: противопожарное опахивание лесных опушек и весеннее выжигание сухой травы на примыкающих к ним луговых склонах, сенокошение (июньское), выпас скота и «вытаптывание растительности» (как правило, свойственно окрестностям населенных пунктов, летним лагерям и зонам отдыха). От этого в первую очередь страдают кормовые растения гусениц (хохлатки) и бабочек (многолетники) [12, 13, 17, 25]. Поскольку в популяциях C. solida, господствует семенное возобновление (вегетативное размножение в самоподдержании популяции данного вида существенной роли не играет), а хохлатка плотная – мирмекохор [19], необходимо организовать охрану некоторых видов муравьев трофически связанных с этим растением. Также нуждаются в охране и основные опылители хохлатки – шмели, причем мелкие их виды в поисках нектара вынуждены прокусывать шпорец цветка (личный сообщение Т. Г. Стойко). Вырубка леса (особенно сплошная) и лесные пожары [26] крайне неблагоприятно сказываются для популяций хохлатки и мнемозины. Отмечены (не редки) случаи гибели бабочек на автомобильных дорогах.
Меры охраны. Мнемозина - один из первых видов среди чешуекрылых насекомых, который был признан нуждающимся в специальных мерах охраны (Goldmann, 1911) [1]. Несмотря на то, что вид включен в большинство Красных книг Европы, в Юго-Западной Финляндии уже вымер P. m. mnemosyne L., близок к вымиранию P. m. osiliensis Viidalepp, 1966 [27]. В Красную книгу РФ (Животные, 2001) - D. mnemosyne (L.) внесена как сокращающийся в численности вид – 2 категория [1]. В КК Пензенской области, Том № 2 (Животные, 2005), вид приведен под 3 категорией – редкий вид. В области охраняется на территории ГПЗ «Приволжская лесостепь» и ряде ООПТ [12].
Благодарности. За постоянную помощь и консультации авторы выражают искреннюю благодарность энтомологам - Ю. П. Коршунову (СО РАН, г. Новосибирск), А. Г. Татаринову (г. Сыктывкар); В. В. Золотухину (УГПУ, г. Ульяновск), Б. В. Страдомскому (Ростов-на-Дону), Ю. Н. Старикову (с. Невежкино, Пензенская обл.) и Л. В. Большакову (г. Тула), В. В. Аникину (СГУ, г. Саратов), С. А. Сачкову (СГУ, г. Самара), А. А. Ластухину (г. Чебоксары), А. В. Свиридову (ЗМ МГУ, г. Москва); зоологам - Т. Г. Стойко, И. В. Муравьеву, Ю. А. Мазею (ПГПУ, г. Пенза); экологам - С. В. Иванову (гимназия № 1, г. Кузнецк) и Л. Н. Ивановой (ПГКМ, г. Пенза); ботаникам - А. А. Солянову, А. А. Чистяковой, Л. А. Новиковой (ПГПУ, г. Пенза), Т. В. Разживиной (ГПЗ «Приволжская лесостепь», г. Пенза); естествоиспытателям и охотникам М. А. Монахову, Р. П. Васьковскому, В. А. Чернышову и Д. А. Капустину. За предоставление своих сборов и дневниковых материалов, членам Пензенского отделения Русского Энтомологического Общества – В. М. Великоиваненко, А. М. Монахову, Е. М. Монахову, Д. В. Поликанину, А. Н. Роганину, А. Е. Барышеву, А. В. Леонтьеву, Д. Н. Подгорнову, В. И. Кузнецову, В. Н. Тремасову и студенткам - Е. В. Заикиной и И. В. Евсиковой.
