

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
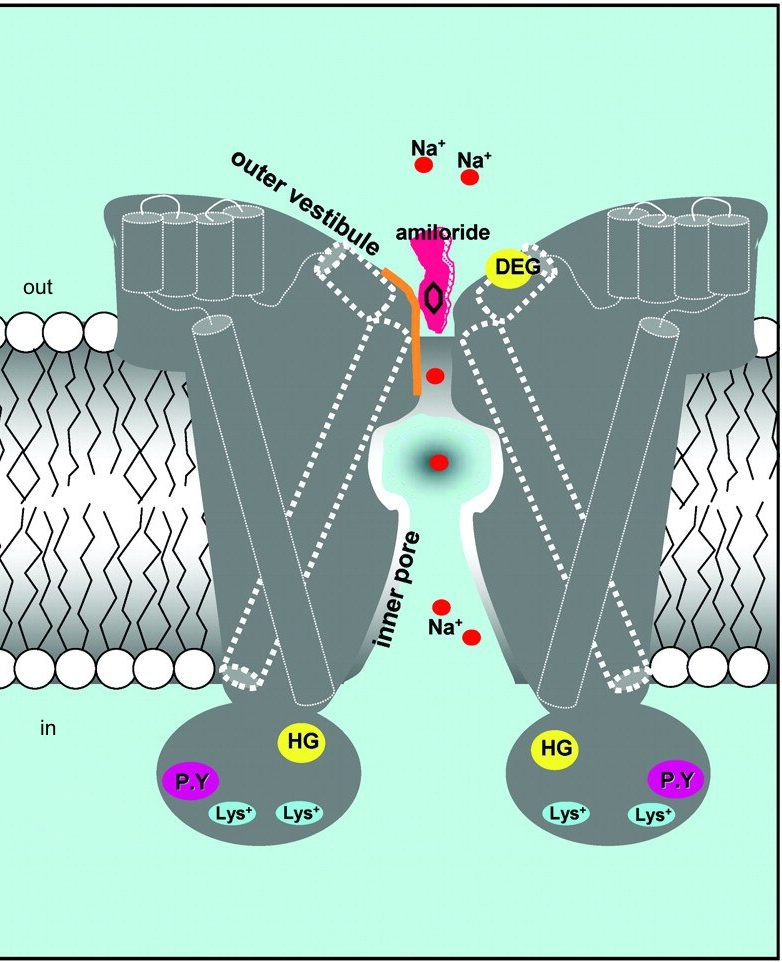
Рис. 9. Модель поры ENaC. (Модифицировано из Loffing, Schild, 2005) |
Селективный фильтр поры ENaC позволяет проникать через канал катионам Na+ и более мелким катионам Li+ и H+, при этом более крупные катионы, такие как катионы K+, Rb+, Cs+, не могут проходить сквозь канал дикого типа (Palmer L. G., 1990). Предполагается, что катионы проходящие через канал полностью лишены гидратной оболочки или связаны не более чем с одной молекулой воды в районе селективного фильтра ( Doyle D. A. et al. , 1998 ). Исходя из радиуса катионов K+ и Na+ было высказано предположение, что диаметр селективного фильтра у ENaC дикого типа составляет 1,9-2,7 Е. На основании исследования селективности каналов с мутациями G587 и S589 в б-субъединице, G529 в в-субъединице (положение гомологично бG587 ) и S542 в г-субъединице (положение гомологично бS589), предполагается, что данный участок непосредственно является селективным фильтром (Snyder P. M. et al., 1999; Kellenberger S. et al., 2001) (Рис. 10).
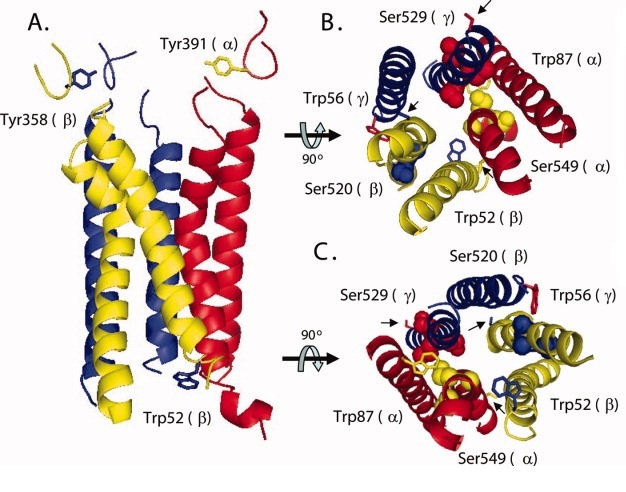
Рис. 10. Предсказанная структура поры и области короны ENaC. Предсказанные порообразующие трансмембранные домены гетеротримерного ENaC параллельно плазматической мембране (А). Консервативный Trp52 на внутриклеточный N-конце TM1 в-hENaC изображен с голубой боковой цепью. Консервативный Tyr (358 и 391) в предполагаемых петлях связи в - и б-ENaC, которые находятся чуть выше поры, изображены с синими и желтыми боковыми цепями, соответственно. Изображение поры, перпендикулярно плазматической мембране с внеклеточной (В) и внутриклеточной (С) стороны, изображены консервативный Trp на N-конце TM1 и консервативный Ser (черные стрелки). На С-конце ТМ2, отмеченном для каждой субъединицы. Боковые цепи остатков в ТМ2 каждой субъединицы, образующей селективный фильтр, размещены на расстоянии, заполненном шариками. (модифицировано из Stockand et al., 2008) |
1.4. Биофизические характеристики ENaC
ENaC являются высокоселективными каналами. Проводимость ENaC для Na+ и Li+ в 100-1000 раз выше, чем для K+. ENaC дикого типа абсолютно непроницаемы для более крупных одновалентных катионов (Cs+, Rb+) и для всех двухвалентных катионов (Palmer L. G. et al., 1982). По-видимому, в связи с меньшими размерами катионов Li+ проводимость ENaC для них выше, чем для Na+ в 1.3-2 раза в зависимости от типа канала и температуры (Kellenberger S. et al., 1999). Диаметр селективного фильтра ENaC дикого типа составляет 1,9-2,7 Е (Kellenberger S. et al., 2001). Проводимость ENaC для Na+ при концентрациях Na+ >100 мМ и комнатной температуре составляет примерно 4-5 пС, в зависимости от типа ENaC и около 9 пС при 37° С (Garty H., Palmer L. G., 1997).
На сегодняшний день выделено несколько типов ENaC. Они различаются по таким характеристикам как: чувствительность к амилориду, селективность, проводимость. Na+-каналы, экспрессирующиеся в дистальных сегментах нефрона млекопитающих и в эпителии кожи и мочевого пузыря амфибий, относятся к типу высоко-селективных или Н-типу ENaC (Garty H., Palmer L. G., 1997; Kellenberger S., Schild L., 2002). Каналы Н-типа ENaC обладают очень высокой чувствительностью к амилориду и максимальной среди ENaC селективностью для катионов Na+ в сотношении к катионам K+ (Benos D. J. et al., 1995; Garty H., Palmer L. G., 1997; Kellenberger S., Schild L., 2002). Проницаемость ENaC каналов Н-типа для ионов Li+ примерно в полтора раза выше, чем для ионов Na+ (Garty H., Palmer L. G., 1997). Помимо этого высокоселективные Na+-каналы обладают самой медленной кинетикой среди всех типов ENaC (Garty H., Palmer L. G., 1997). Средняя длительность открытого и закрытого состояния канала составляет (при комнатной температуре) 0,5 – 5 с (Garty H., Palmer L. G., 1997; Kellenberger S., Schild L., 2002, и др., 2006).
ENaC блокируются пиразинкарбоксиамидом — амилоридом в микромолярной концентрации (Bentley P. J., 1968; Benos D. J. et al., 1976) (Рис. 11). Амилорид был разработан в 1960-х годах, как K+-сберегающий диуретик (Baer J. E. et al., 1967). Блокирование ENaC амилоридом является потенциал-зависимым и усиливается при гиперполяризации мембраны (Palmer L. G., 1984; Garty H., Palmer L. G., 1997). Было установлено, что мутации в бSer583, вGly525 и гGly537 снижают чувствительность ENaC к амилориду. Также было показано, что снижение чувствительтности к амилориду зависит от того, в какой субъединице происходит мутация и какая аминокислотная замена происходит (Schild L. et al., 1997; Kashlan O. B. et al., 2005). С помощью поли - и моноклональных антител к амилориду в CRD I была обнаружена последовательность WYRFHY, мутации в которой приводили к уменьшению амилорид-чувствительности (Kieber-Emmons T. et al., 1995; Li X. J. et al., 1995). Однако роль данного участка в блокировании канала не ясна.
ENaC обладают саморегуляцией. Катионы Na+ ингибируют ENaC двумя путями. Во-первых, так называемое, Na+-ингибирование обратной связи. Данный процесс вызывается ростом внутриклеточной концентрации Na+ и приводит к уменьшению активности ENaC. Такая инактивация развивается в течение нескольких минут и связанa с уменьшением числа активных каналов на поверхности клеток и уменьшением вероятности открытого состояния канала (Frindt G. et al., 1993, 1995; Anantharam A. et al., 2006). Во-вторых, Na+-самоингибирование, вызывающее быстрое снижение активности ENaC после роста концентрации катионов Na+ во внеклеточной среде (Chalfant M. L. et al., 1999; Horisberger J. D., Chraпbi A., 2004; Bize V., Horisberger J. D., 2007). Данные процессы направлены на уменьшение входа Na+ на одном участке, с целью оптимального поглащения вдоль всего дистального нефрона (Kellenberger S., Schild L., 2015).
Недавно было показано, что анионы Cl - ингибирует ENaC. Анионы Cl - ингибируют ENaC за счет уменьшения вероятности открытого состояния канала. При этом Br - и I - ингибируют ENaC аналогично Cl-, а F - и более крупные фосфаты и сульфаты ингибируют ENaC очень слабо (Collier D. M., Snyder P. M., 2009, 2010).
Ввиду структурной общности и нахождению в одном суперсемействе с дегенеринами, было предположено, что ENaC обладают механочувствительностью. Данное предположение было подтверждено экспериментами. Считается, что на активацию ENaC влияет поток жидкости, каким-то образом изменяющий конфигурацию экстраклеточных доменов. Полагается, что с механочувствительностью каким-то образом связаны CRD-домены (Tavernarakis N., Driscoll M., 2000; Kellenberger S., Schild L., 2002; Carattino M. D. et al., 2003) (Рис. 12). Обнаружена экспрессия в - и г-субъединиц ENaC в механо-чувствительных нейронах, а также установленно, что амилорид и бензамил (блокаторы ENaC) препятствуют передаче механического стимула в барорецепторных нейронах (Drummond H. A. et al., 2001). Отсутствие экспрессии б-субъединицы, позволяет предположить, что активный канал в нейрональных клетках не образуется, однако остальных двух субъединиц, возможно, достаточно для выполнения роли механозависимых сенсоров (, , 2009).
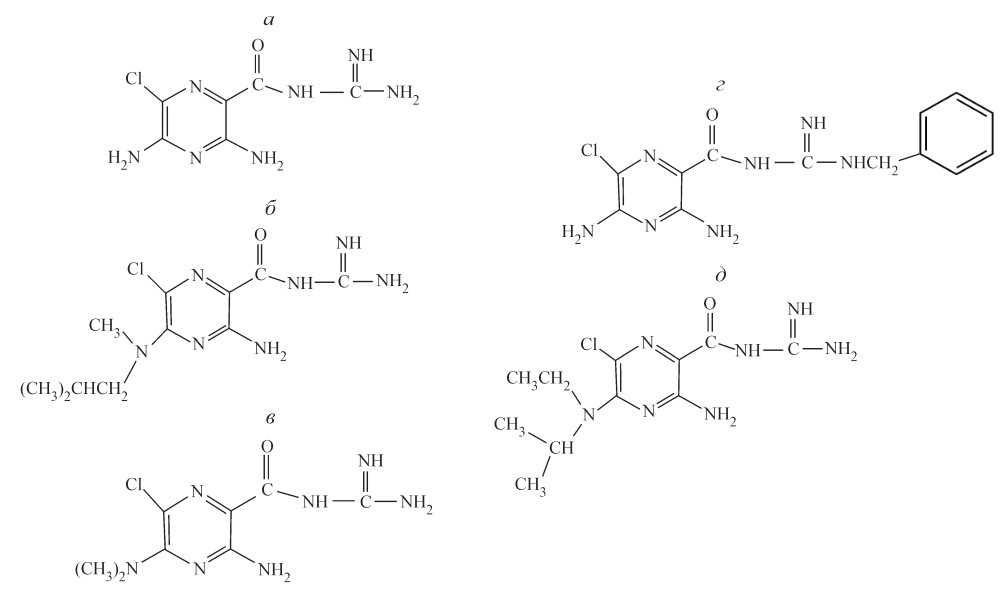
Рис. 11. Блокаторы ENaC. А – амилорид (3,5-диамино-N-(аминоиминометил)-6-хлорпиразинкарбоксамид). Б — метил изобутил амилорид. В — диметил амилорид. Г — бензамил. Д — 5-(N-этил-N-изопропил)-амилорид. (Модифицировано из Мельницкая и др., 2006) |
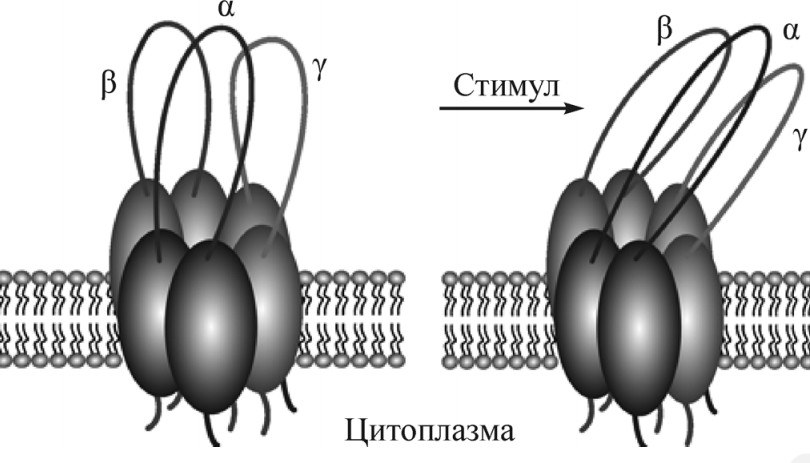
Рис. 12. Возможная модель механочувствительности ENaC. Внеклеточные петли спокойно находятся с внешней стороны клеточной мембраны (слева). Реакция на изменения скорости и давления внеклеточной жилкости. (Модифицировано из Вачугова, Морачевская, 2009) |
2. Функция ENaC в различных тканях и органах
2.1. Функция ENaC в почках
Почки поддерживают постоянство состава внеклеточной жидкости (Рис. 13). Почки выводят из организма избыток воды, растворенные в ней вещества. Наоборот, при дефиците воды и электролитов включаются процессы, уменьшающие их дальнейшие потери, сохраняя нормальную экскрецию продуктов обмена. Функциональной единицей почки является нефрон. Нефрон состоит из клубочка с боуменовой капсулой, проксимального извитого канальца, петли Генле, дистального извитого канальца и собирательной трубочки.
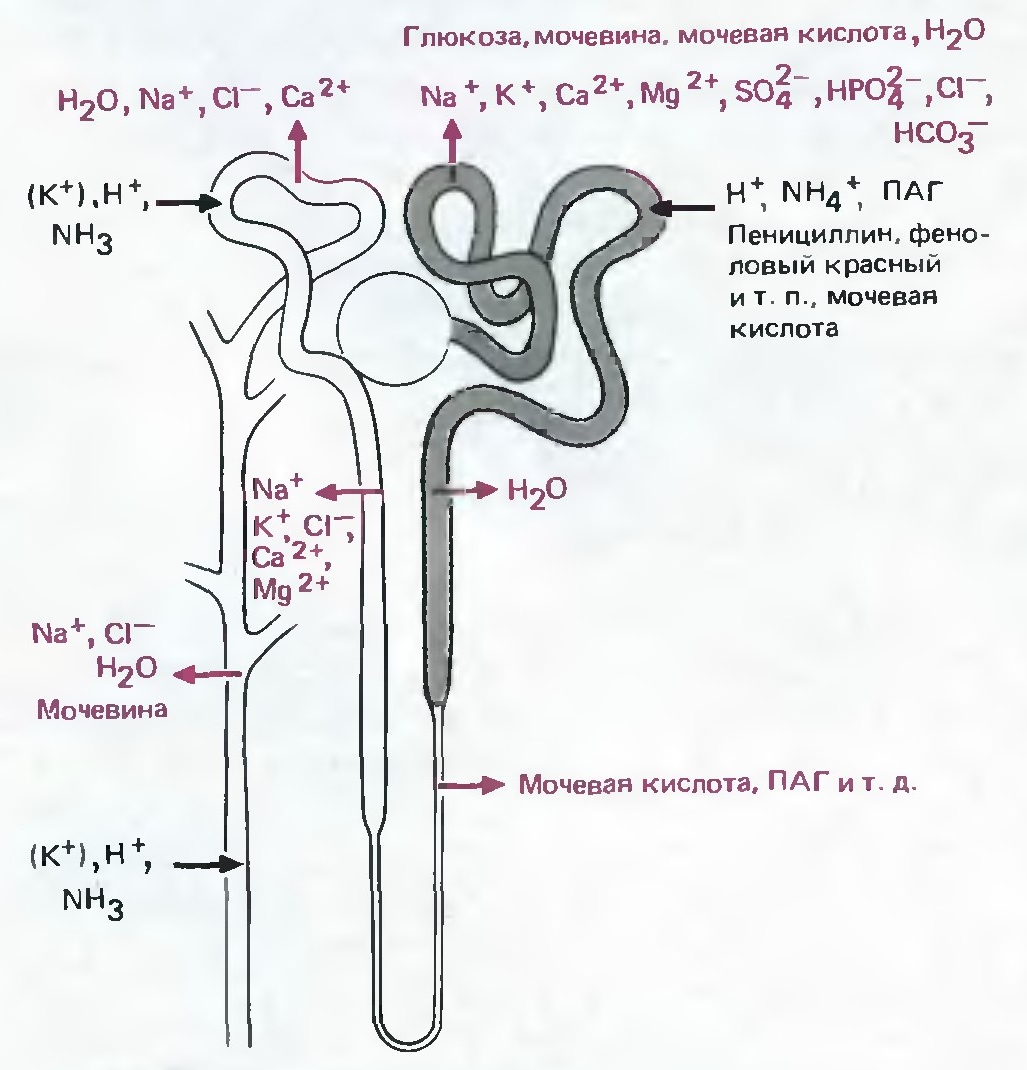
Рис. 13. Локализация различных транспортных процессов в нефроне. Красным отмечены реабсорбируемые вещества. Черным — секретируемые. (Модифицировано из Шмидт, Тевс, 2005) |
Почка является основным органом, регулирующим экскрецию Na+. Обычно, в почке реабсорбируется до 99% процентов Na+, отфильтрованного в капиллярном клубочке (Рис. 14). Почки способны очень тонко контролировать количество экскретируемого Na+. 60-70% Na+ реабсорбируются в проксимальном извитом канальце, 15-25% - в петле Генле, 5-10% - в дистальном извитом канальце и 1-2% в собирательной трубочке. В реабсорбции Na+ в почке участвуют такие транспортные белки как Na+/H+-обменник, в проксимальном извитом канальце, Na+/K+/Cl-—ко-транспортер в петле Генле, Na+/Cl--ко-транспортер в дистальном извитом канальце, ENaC, Na+/K+-АТФаза, снабжающая энергией процесс активного переноса Na+ (Sahay M. et al., 2007) (Рис. 15).
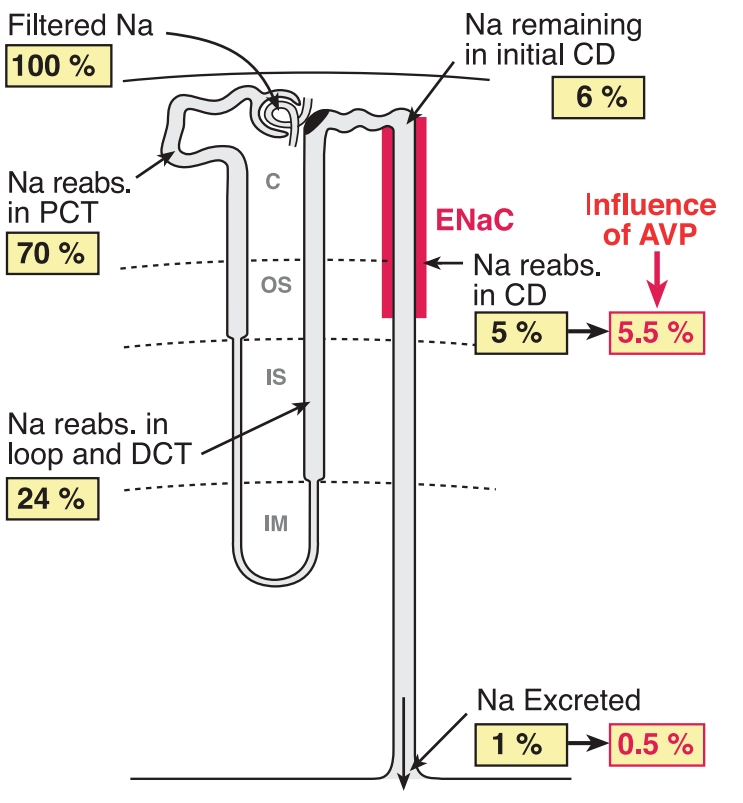
Рис. 14. Количество Na+ реабсорбированого в нефроне. CD – собирательная трубочка. DCT – дистальный извитой каналец. PCT – проксимальный извитой каналец. (Модифицировано из Bankir et al., 2010) |
В почке ENaC выполняет функцию тонкой настройки уровня экскреции Na+, поддержания K+ гомеостаза, корректировку кислотно-щелочного баланса. Экспрессируется ENaC в почке, в основном, в собирательных трубочках, а также в дистальных извитых канальцах (Rossier et al., 2013) (Рис. 15). В тех же клетках экспрессируются V2-рецептор и аквапорин-2. Экспрессия ENaC регулируется транскрипционными и пост-трансляционными факторами. Эта регуляция происходит непосредственно на субъединицах канала и/или на ENaC-регулирующих белках (Loffing J. et al., 2001) Регуляция ENaC в почке происходит под действием гормонов: вазопрессина и альдостерона (Loffing J. et al., 2000). Принцип действия этих гормонов, в основном, заключается в увеличении плотности ENaC на апикальной мембране клеток (Butterworth M. B. et al., 2009). Вазопрессин увеличивает плотность ENaC за счет слияния везикул содержащих ENaC с апикальной мембраной (Butterworth M. B. et al., 2005). После удаления вазопрессина, ENaC подвергается эндоцитозу с поверхности мембраны и реорганизуется в рециркулирующие везикулы. Альдостерон может регулировать активность ENaC с помощью транскрипционно-зависимых и независимых механизмов (Thomas et al., 2007). Например, альдостерон увеличивает плотность ENaC за счет уменьшения интернализации ENaC, путем синтеза киназы SGK1, которая негативно модулирует Nedd4 (Debonneville C. et al., 2001). Нарушение структурно-функциональной организации ENaC приводит к тяжелым наследственным заболеваниям (синдром Лиддла, псевдогипоальдостеронизм I типа и др.).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
