
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Преимущества Моей Технологии Клонирования
Все широко используемые методы клонирования ДНК в плазмиды чудовищно неэффективны – хотя из рекламы производителей китов для клонирования вы об этом вряд ли узнаете. Дело в том, что «эффективностью клонирования» обычно называется доля колоний с инсертами (отношение количества колоний с плазмидами с желаемыми инсертами к общему количеству колоний). При бело-голубой селекции, есть очень похожее понятие фона, только со знаком минус – это доля голубых колоний без инсертов. Эффективностью же клонирования следовало бы называть нечто совсем другое; но не будем переопределять устоявшийся термин. Введем новый: КПД клонирования, связанный с эффективностью трансформации бактерий суперскрученными плазмидами.
Пусть, для примера, эффективность трансформации химически компетентных бактерий равна 10⁹ cfu/мg суперскрученной pUC19. Разумеется, целый микрограмм плазмиды для этого никто не использует – используют порядка 1 пикограмма pUC19 (или меньше). Это даст примерно 1000 колоний на чашку Петри, что уже можно посчитать. При этом, 1 пикограмм pUC19 (2686 bp) = 0.57 аттомолей = 3.4‧10⁵ молекул. Теперь зададим вопрос: сколько колоний с инсертами можно получить, если использовать для клонирования эквимолярное количество молекул фрагментов ДНК (т. е. 0.57 аттомолей)? Ответ: ноль. Чтобы получить сравнимое (т. е. порядка 1000) число колоний с инсертами, необходимо использовать в десятки и сотни тысяч раз большее число молекул фрагментов ДНК, чем число суперскрученных плазмид при проверке эффективности трансформации.
Пусть перед клонированием было N молекул фрагментов ДНК. После клонирования, получилось М колоний бактерий, содержащих плазмиды с инсертами. Если же взять эквимолярное фрагментам ДНК количество суперскрученных плазмид (N молекул) и трансформировать ими бактерии, то получится К колоний. Определим КПД клонирования как отношение числа колоний с инсертами к числу колоний, продуцируемых количеством суперскрученной плазмиды, эквимолярным количеству фрагментов ДНК до клонирования (N): КПД = М/К. Как правило, КПД клонирования значительно меньше 0.1%.
Другие проблемы всех методов клонирования:
• Чем длиннее ДНК, тем хуже она клонируется и тем чаще подвергается делециям. Фрагменты ДНК длиной 10 kb имеют КПД клонирования в десятки раз ниже 1 kb фрагментов, 20 kb фрагменты – в сотни раз ниже 10 kb фрагментов. Результатом такой сильнейшей селекции на короткие инсерты является то, что библиотеки ДНК (скажем, кДНК) всегда сильно обеднены длинными фрагментами ДНК.
• Часть колоний содержит плазмиды без инсертов и с делециями инсертов или участков самой плазмиды. Важно, что чем меньше количество молекул фрагментов ДНК до клонирования, тем выше процент плазмид без инсертов и с делециями. В результате, малые количества ДНК клонировать почти невозможно – единичные колонии с инсертами теряются на фоне большого числа колоний без инсертов и с делециями. Бело-голубая селекция и клонирование в токсичные гены не решают эту проблему – делеции инсертов и участков плазмиды дают большое количество фальшивых позитивов.
• Определенные последовательности ДНК чрезвычайно трудно клонировать. Таковы, например, прямые и инвертированные повторы (палиндромы), теломеры, центромеры. Как результат, 8% генома человека до сих пор не клонированы и не секвенированы. Еще хуже обстоят дела с геномами животных и растений. Это относится и ко многим кДНК, содержащим повторы и палиндромы.
Изобретенная мной технология клонирования решает все эти проблемы – причем со 100% КПД и эффективностью:
• КПД клонирования равен 100% - то есть, например, 1000 молекул фрагментов ДНК с тупыми концами продуцирует такое же число колоний с инсертами, как и 1000 молекул суперскрученной плазмиды.
• Эффективность клонирования равна 100% - то есть плазмиды без инсертов или с делециями инсертов и участков самой плазмиды полностью отсутствуют.
• Это позволяет клонировать очень малые количества ДНК (единицы фемтограмм), а также геномные ДНК и кДНК из отдельных клеток.
• Селекция на короткие инсерты отсутствует – то есть и короткие, и длинные фрагменты ДНК клонируются с равными эффективностью и КПД. Для примера, смесь эквимолярных количеств 1 kb и 10 kb ПЦР продуктов после клонирования дает равное число колоний с 1 kb и 10 kb инсертами; плазмиды без инсертов и с делециями отсутствуют.
• Технология позволяет клонировать фемтограммные количества очень длинных продуктов ПЦР (десятки kb) после всего 3 (трёх) циклов ПЦР. На агарозном геле их не видно, но они клонируются.
• Очень длинные фрагменты геномной ДНК (сотни kb) клонируются в плазмиды (с низким числом копий на клетку) с КПД, всего в несколько раз меньшим, чем у коротких фрагментов.
• Повторы, палиндромы, теломеры, центромеры клонируются со 100% эффективностью и КПД, как и любые другие фрагменты ДНК.
• Это позволяет – впервые – создать полностью репрезентативные геномные и кДНК библиотеки, полностью клонировать и секвенировать геномы и кДНК любых организмов.
Частично эта технология клонирования описана в US Patent 6825011 и EPO Patent 1141239-A1. Технология была многократно проверена в экспериментах, оптимизирована и коммерциализирована.
С 1992 г., я работал в США, сначала в University of Texas Southwestern Medical Center at Dallas и Memorial Sloan-Kettering Cancer Center, New York. Затем я стал одним из основателей и главным ученым Gene Source, Inc., в основу деятельности которой легли изобретенные мной технологии. В 2000 г. мы продали эксклюзивную лицензию на US Patent 6825011 Life Technologies, Inc., которая применяла ее для клонирования продуктов ПЦР, создания геномных и кДНК библиотек. Однако, вскоре Life Technologies была куплена своим главным конкурентом – Invitrogen, который уволил практически всех исследователей и закрыл лаборатории своего бывшего конкурента. Invitrogen не стал инвестировать в коммерциализацию моей технологии (они решили, что им достаточно TOPO Cloning), несколько лет они платили нашей компании minimum royalties, затем лицензия была прекращена.
Поскольку эта технология была полностью апробирована и оптимизирована, мы решили заняться ее коммерциализацией сами, на собственные деньги. Одновременно, мы занимались разработкой массово-параллельной технологии синтеза ДНК (до нескольких сотен тысяч пар оснований) и ДНК-микрочипов очень высокой плотности (несколько миллионов проб на смІ). Но бизнес не наука – денег нам не хватило и в конечном итоге наша компания обанкротилась, несмотря на то, что наши технологии в тысячи раз более эффективны, чем все альтернативные методы. Однако, банкротство компании не означает конец ее технологий.
Сейчас я вернулся в Россию и ищу компанию или организацию, которая нуждается в клонировании “неклонируемых” ДНК и создании действительно репрезентативных геномных и кДНК библиотек. Эта технология клонирования позволяет также создать массово-параллельную технологию синтеза ДНК со стоимостью синтеза 1 пары оснований в тысячи раз меньшей, чем у других коммерчески доступных методов. Сейчас ваша компания или организация имеет шанс стать единственным бенефициаром этих технологий, если я буду применять их у вас.
Экспериментально доказать, что моя технология клонирования действительно в тысячи раз превосходит все остальные методы, я могу в течение 2 месяцев. Достаточно для испытательного срока?
На период демонстрации, меня устроит позиция научного сотрудника. После демонстрации, позиция руководителя лаборатории/направления позволит мне применить технологии клонирования и синтеза ДНК в более широких масштабах. Насколько широких? Чтобы в конечном итоге компания стала мировым лидером в клонировании и синтезе ДНК. Это не преувеличение – но на это нужен достаточный уровень финансирования, получить которое можно, например, через государственные гранты и инвестиционные фонды. Я ожидаю, что руководство компании имеет достаточные связи и опыт получения грантов/инвестиций.
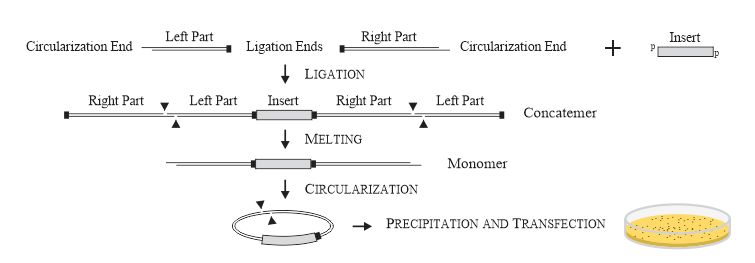
Суть патентованной технологии
Клонирование делится на несколько этапов, эффективность каждого из которых близка к 100%:
1. Фосфорилированные фрагменты ДНК смешиваются с многократным избытком дефосфорилированной плазмиды, состоящей из двух частей с длинными комплементарными липкими концами, и ДНК-лигазой в буфере, содержащем 10% PEG 6000. Время инкубации – 5 мин. Эффективность лигирования – 100%. В этих условиях, все комплементарные липкие концы плазмид прилипают друг к другу.
2. Объем увеличивается в несколько раз и смесь молекул нагревается на 2 мин до такой температуры, что все длинные липкие концы «плавятся». Эффективность плавления – 100%. Каждый инсерт в этой момент оказывается частью линейного комплекса: левая половина плазмиды – инсерт – правая половина плазмиды.
3. Смесь охлаждается до температуры, при которой длинные липкие концы комплементарно взаимодействуют друг с другом. Время инкубации – 5 мин для инсертов короче 10 kb, 2 часа для инсертов несколько сотен kb длиной. Все комплексы левая половина плазмиды – инсерт – правая половина плазмиды циркуляризируются (замыкаются в кольца). Эффективность циркуляризации – 100%.
4. Смесь смешивается с компетентными клетками и производится трансформация. Эффективность трансформации – 100%, то есть такая же, как у эквимолярного количества суперскрученной плазмиды.
Подчеркну, что ряд важнейших ноу-хау не включен в патент. Без этих ноу-хау, избежать делеций инсертов и частей самой плазмиды невозможно, а как следствие, невозможно клонировать палиндромы, теломеры, центромеры.
Юрий Михайлович Романчиков, к. б.н.
Тула
Тел: +7-953-961-7874
yury. *****@***com
ЦЕЛЬ
• Использовать патентованную технологию клонирования в компании, которая нуждается в клонировании “неклонируемых” ДНК и создании действительно репрезентативных геномных и кДНК библиотек, включая повторы, палиндромы, теломеры, центромеры.
РЕЗЮМЕ
• Главный Ученый и один из основателей Gene Source Inc. (США), специализирующейся в клонировании и синтезе ДНК.
• Перед этим, работал в ведущих научных учреждениях России и США, изучая молекулярные механизмы старения и рака.
• Изобретатель широкого спектра технологий, в том числе - клонирования и синтеза ДНК, ДНК-микрочипов высокой плотности.
• Автор 4 теорий онтогенеза, старения и рака, теории происхождения жизни на Земле, ряда физических теорий.
• Инженер-электрофизик (ФТФ НЭТИ), к. б.н. (Институт Биологии Развития РАН).
ДОСТИЖЕНИЯ
• Изобрел технологию клонирования ДНК с эффективностью более чем в 1000 раз выше, чем у всех других методов.
• Изобрел массово-параллельную технологию синтеза ДНК до нескольких сотен тысяч пар оснований, позволяющую уменьшить стоимость синтеза ДНК более чем в 1000 раз.
• Изобрел массово-параллельную технологию создания ДНК-микрочипов очень высокой плотности.
• Изобрел 2 метода быстрого субклонирования из одной плазмиды в другую.
• Изобрел метод синтеза кДНК полной длины.
• Изобрел метод преимущественного клонирования длинных последовательностей ДНК.
• Изобрел концентраторы биохимических и химических веществ без физической и химической обработки.
• Изобрел технологию очень дешевого опреснения морской воды.
• Клонировал главный ген-мишень мышиного фактора транскрипции E2F и 5 генов-мишеней суппрессора опухолей р53 из человеческого и мышиного геномов.
• Изобрел метод искусственного удлинения теломер без помощи теломеразы в нормальных клетках животных.
• Автор 4 альтернативных теорий онтогенеза, старения и рака.
• Автор теории происхождения жизни на Земле в горячих и гидрофобных условиях.
• Автор ряда физических теорий.
ДОСТОИНСТВА И КВАЛИФИКАЦИИ
• Теоретик, экспериментатор, изобретатель, аналитик • Уникальная способность решения сложных проблем • Честный, дружелюбный, надежный, неконфликтный • Большой опыт руководства коллективом, обучения сотрудников, патентования, ведения бизнеса в США
• Клонирование “неклонируемых” ДНК • Быстрое и эффективное субклонирование
• Создание репрезентативных геномных и кДНК библиотек • Секвенирование • Синтез кДНК полной длины • PCR, RT-PCR и qRT-PCR • Сайт-направленный мутагенез • EMSA
• Дифференциальный дисплей • Northern, Southern и Western блоттинг • In vitro транскрипция и трансляция • Электрофорез ДНК, РНК, белков • HPLC • Гель-фильтрация • ELISA
• Иммунопреципитация • Иммунофлуоресценция • Клеточные культуры млекопитающих и бактерий • ДНК-микрочипы • Селекция кандидатов в лекарства • Онкогены и опухолевые суппрессоры • Теломеры и теломераза • Программы для работы с ДНК, РНК и белками
• Mathcad • Fortran • HTML
ПРОФЕССИОНАЛЬНЫЙ ОПЫТ
Gene Source Inc., Germantown, MD........................................................Май 1997 – Март 2017
Chief Scientific Officer (Главный Ученый)
Изобрел технологии, которые легли в основу научной деятельности Gene Source Inc. Руководил развитием, патентованием, лицензированием и коммерциализацией этих технологий.
Memorial Sloan-Kettering Cancer Center, New York, NY.........................Aug 1994 – Apr 1997
Research Fellow (Научный Сотрудник)
Изобрел метод клонирования генов-мишеней факторов транскрипции. Используя этот метод:
• Клонировал главный ген-мишень мышиного фактора транскрипции E2F, который играет ключевую роль в регуляции клеточных делений.
• Клонировал 5 генов-мишеней суппрессора опухолей р53 из человеческого и мышиного геномов.
• Доказал, что зависимая от клеточного цикла транскрипция гена мышиного гистона Н3 регулируется несколькими E2F сайтами на его 3’ конце.
University of Texas Southwestern Medical Center, Dallas, TX.........Июнь 1992 – Август 1994
Research Fellow (Научный Сотрудник)
• Изобрел метод искусственного удлинения теломер без помощи теломеразы в нормальных клетках животных (путем экспрессии обратной транскриптазы и РНК с повторами CCCUAA).
• Определил ДНК-связывающие сайты для нотивных и мутантных белков р53, циклинов и циклин-зависимых киназ, используя метод селекции на основе ПЦР (CASTing).
Институт Биологии Гена РАН, Москва........................................Октябрь 1991 – Июнь 1992
Научный Сотрудник
• Участвовал в клонировании и секвенировании 2 генов Xenopus.
Всесоюзный Онкологический Научный Центр, Москва......Январь 1989 – Октябрь 1991
Младший Научный Сотрудник
• Автор 4 альтернативных теорий онтогенеза, старения и рака, опирающихся на факт, что гены, ответственные за старение и иммортализацию клеток, являются регуляторами активности генов.
• Руководил группой, занимающейся экспериментальной проверкой этих теорий.
ОБРАЗОВАНИЕ
К. б.н. (1992) Диссертация: “Теоретический анализ проблемы старения и иммортализации клеток животных: Возможная роль ядерных протоонкогенов”.
Защита в Институте Биологии Развития РАН, Москва.
Инженер-электрофизик (1982). Физико-технический факультет Новосибирского электротехнического института, Новосибирск.
ПАТЕНТЫ
Romantchikov Yuri: Methods for insertion of nucleic acids into circular vectors. US Patent 6825011.
Romantchikov Yuri: Improved methods for insertion of nucleic acids into circular vectors. EPO Patent 1141239-A1.
Romantchikov Yuri: Improved methods for insertion of nucleic acids into circular vectors. PCT Patent WO/2000/036088.
ПУБЛИКАЦИИ
1. 1992. Ключевая Роль Ядерных Прото-Онкогенов в Старении и Иммортализации Клеток Животных. Известия Российской Академии Наук, Сер. Биол., No 4, 985-988.
2. 1991. Возможные Механизмы Действия Факторов Роста и Старение Клеток. Генетика 27, No 11, 1369-1377.
3. 1991. Триггерный Подход к Пролиферации, Старению и Иммортализации Клеток Животных. Генетика 27, No 10, 1223-1230.
4. 1991. Гипотеза об Экстрахромосомных Регуляторных Генах. Успехи Соврем. Биологии 111, No 5, 643-653.
5. 1991. Онтогенез животных с позиции отсутствия самоподдержания синтеза регуляторных белков. Генетика 27, No 10, 1214-1222.
6. 1991. Факторы Роста, Вторичные Мессенджеры и Онкогены. Успехи Соврем. Биологии 111, No 1, 19-33.
7. 1988. Иерархия Регулонов: Связь с Делением и Дифференцировкой Клеток. Успехи Соврем. Биологии 105, No 3, 406-422.
