

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Некоторые участки спирали могут отличаться по параметрам от α-спирали. Это связано с наличием в полипептидной цепи некоторых аминокислотных остатков - Пролина или Глицина, которые не могут образовывать Н-связи с соответствующими кислородами. Стабилизация таких участков осуществляется за счет образования дисульфидных мостиков, возникающих между аминокислотными остатками (Цистеин и Метионин). Сближение двух SH-групп приводит к окислению - потере атома Н. Возможны связи между радикалами, содержащими ОН - и NH-группы. (рисунок α-спирали).
Второй основной тип вторичной структуры полипептидной цепи - β-цепь или β-шпильки. Исторически, правда, именно эта структура белка была первой описана Астбюри в 1941 г. на основании РСА-исследований β-кератина. Через 10 лет Полинг и Кори установили, что β-структура, или «складчатый лист» – это стабилизированный межцепочечными Н-связями ассоциат вытянутых, зигзагообразных цепей. В зависимости от взаимной ориентации цепей различают параллельные и антипараллельные β-структуры. При этом образуются не продольные водородные связи между пептидными группами, как в случае α-спирали, а поперечные Н-связи. (рисунки параллельной и антипараллельной β-цепи). Края антипараллельных β-структур образованы особым видом вторичной структуры, который называется β-изгибом (реверсионным поворотом). β-изгибы образуются 4 последовательно расположенными амионкислотными остатками, как правило, образующими Н-связи 4→1. Анализ показывает. Что возможны 2 основных типа β-изгибов (так называемые изгибы I и II типа (показать рисунок), отличающиеся ориентацией пептидного карбонила по отношению к средней плоскости 10-членного цикла. β-изгибы – характерный элемент пространственной структуры природных и синтетических олигопептидов, как линейных, так и циклических. Фрагмент β-структуры из двух антипараллельных цепей с β-изгибом часто называют «β-шпилькой».
Теперь рассмотрим регулярные структуры без водородных связей внутри каждой, но соединенные водородными связями между собой.
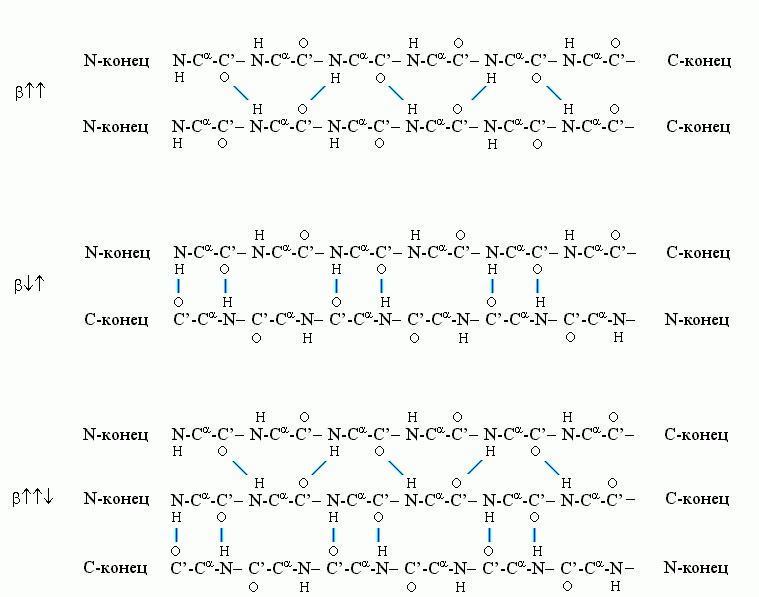
Рис.7-6. Схема хода цепи и расположения водородных связей в параллельной β, антипараллельной β и смешанной β-структуре. Видно, что Н-связи одного остатка в β-тяже направлены одну сторону, следующего - в противоположную, и т. д.
β-структуру образуют почти вытянутые слегка скрученные цепи (все углы в главной цепи - почти trans). Она бывает (Рис.7-6) параллельной, антипараллельной и смешанной.
β-структура стянута водородными связями (символы / , \ и | на схемах 7-6). Она существует в виде более или менее крупных листов. Так как поверхность β структуры - рифленая, ее еще называют "складчатой β-структурой" (Рис.7-7).
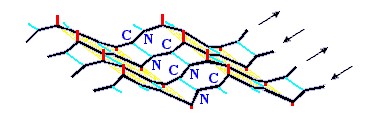
Рис.7-7. Лист β структуры имеет складчатую поверхность. Боковые группы (маленькие отростки) расположены на складках; каждая обращена в ту же, что и складка, сторону, т. е. направленные вниз и вверх боковые группы чередуются вдоль β-тяжа.
β-структурные листы всегда несколько скручены (Рис.7-8а) из-за того, что несколько скручены (Рис.7-8б) отдельные β-тяжи, и потому по ходу тяжа несколько меняется направленность водородных связей. А тяжи, в свою очередь, скручены из-за того, что наиболее энергетически выгодная конформация остатков с боковыми группами сдвинута к центру стерически разрешенной области (Рис.7-8в). Скрученность отдельного β-тяжа - левая (у L аминокислот! - у D было бы наоборот): вы видите (Рис.7-8б), что боковые группы тяжа поворачиваются по часовой стрелке (на ≈-165o на каждый остаток) по мере приближения тяжа к нам.
В результате, поворачиваются и Н-связи (на ≈-165o на каждый остаток, т. е. на -330o=+30o на пару остатков - повторяющуюся единицу β-структуры), так что угол между соседними тяжами β-структуры (если смотреть с кромки листа, Рис.7-8а) обычно составляет около -25о ("минус", как всегда, означает, что ближний к нам β-тяж повернут по часовой стрелки относительно более далекого). Таким образом, β-лист имеет левое скручивание, если смотреть с края этого листа (и правое, если смотреть вдоль β-тяжей на поворот линии Н-связей).
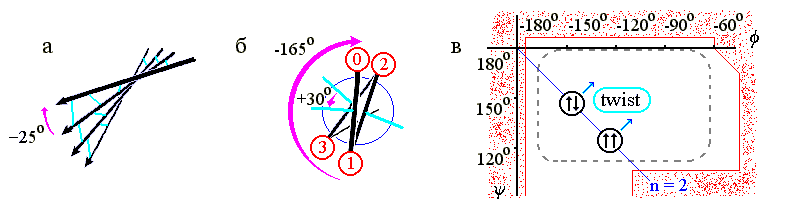
Рис.7-8. (а) Скрученность β-листа. β-тяжи изображены стрелками, водородные связи между ними - голубыми линиями. (б) Схематическое изображение одного витка β-тяжа, вид с торца. Кружки - боковые группы; их номера возрастают по мере удаления от читателя. Голубые линии укаывают направление С=О групп, завязывающих Н-связи в листе, большей стрелкой - поворот β-тяжа при приближении к нам на один остаток, меньшей стрелкой - поворот направленных в одну сторону водородных связей при приближении к нам на два остатка. (в) Конформация идеальной (не скрученной) параллельной и антипараллельной β-структуры для поли(Gly), и усредненная конформация реальной (сложенной из L аминокислот) скрученной (twist) β-структуры. Пунктир показывает область энергетического минимума для отдельно взятого остатка Ala; контуром показаны границы области разрешенных конформаций этого остатка. Диагональ φψ-карты соответствует плоской периодичной структуре, имеющей 2 остатка на виток. Над диагональю лежат левые (L) спирали, под ней - правые (R).
Третий тип спирали принято называть пролиновыми спиралями. Это спирали полипролин I и полипролин II. Первая из них – левая спираль, содержащая 10 остатков пролина на 3 витка. Все пептидные связи в ней находятся в цис-конформации, что редко встречается в обычных полипептидных структурах. Вторая полипролиновая спираль – левая, с пептидными связями в транс-положении. В нормальных физиологических условиях более стабильна вторая спираль. Она имеет 3 остатка на виток и смещение вдоль оси спирали 3.12Е на остаток по сравнению с 1.5Е для α-спирали. Такая полипролиновая спираль имеет более вытянутую цепь, чем α-спираль.
Есть спирали и без водородных связей, где плотная (а значит, энергетически выгодная) упаковка держится чисто на Вандерваальсовых контактах. Это полипролиновая спираль. При этом три скрученные в довольно растянутую левую спираль цепи образуют правую суперсуперспираль - они плотно закручиваются друг вокруг друга. Из двух возможных типов полипролиновой спирали для нас важна спираль poly(Pro)II: она реализуется в коллагене. В этой спирали пептидные группы пролинов находятся в обычной (trans) конформации. Отложим более подробное рассмотрение коллагеновой спирали до соответствующего места курса, а пока ограничимся общим ее видом (Рис.7-9) и отметим на Рис.7-4 область соответствующей ей конформации цепи: видно, что она довольно близка к β структуре.

Рис.7-9. Общий вид тройной правой суперспирали из левых спиралей Poly(Pro)II.
Параметры наиболее важных регулярных вторичных структур белковых цепей суммируются в следующей таблице:
Основные геометрические параметры наиболее распространенных в белках вторичных структур.
Структура | H-связь | Остаток/виток | Смещение/остаток (Е) | ϕ | ψ |
Спираль αR | CO0—HN+4 | +3.6 | 1.5 | -600 | -450 |
Спираль (310)R | CO0—HN+3 | +3 | 2.0 | -500 | -250 |
Лист β↓ | меж цепей* | -2.3 | 3.4 | -1350 | +1500 |
Лист β | меж цепей* | -2.3 | 3.2 | -1200 | +1350 |
Спираль Poly(Pro) II | нет | -3 | 3.0 | -800 | +1550 |
* Расстояние между тяжами в β-листе: 4.8 Все цифры округлены. "+" означает правую спираль, "-" - левую.
Кроме регулярных, в полипептидных цепях есть еще и нерегулярные вторичные структуры, т. е. стандартные структуры, не образующие длинных периодических систем. Это так называемые β-изгибы ("β" - потому, что они часто стягивают верхушки соседних β-тяжей в антипараллельных β шпильках). Характерный вид наиболее важных β-изгибов и конформации входящих в них остатков представлены на Рис.7-10. Сравните Рис.7-10в с Рис.7-4 и Таблицей 7/1, и обратите внимание на то, что изгибы I (и особенно III) близки по конформации к витку спирали 310.
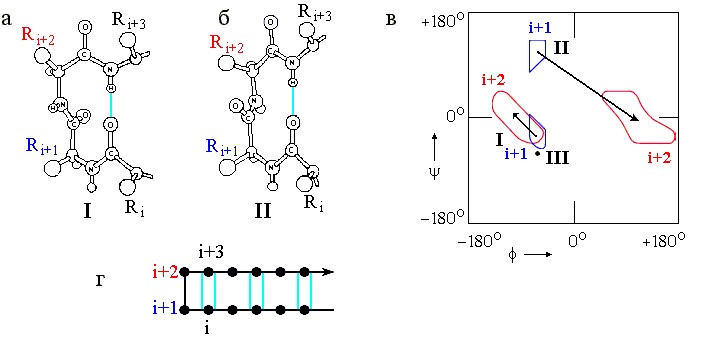
Рис.7-10. β-изгибы. (а) β-изгиб типа I (β-изгиб типа III очень на него похож и потому не нарисован отдельно). (б) β-изгиб типа II. Его основное отличие от β-изгиба I - переворот пептидной группы, соединяющей остатки i+1 и i+2. (в) Конформации фиксируемых водородной связью остатков i+1 и i+2 в β-изгибах. В β-изгибе III оба остатка i+1 и i+2 имеют одинаковую конформацию (отмечена жирной точкой). Конформации остатков i и i+3 в β-изгибах не фиксированы; они фиксируются β-структурой - если она прирастает из изгиба, как на рисунке (г), где дана схема β-шпильки с β-изгибом в ее вершине.
Теперь несколько слов о том, как экспериментально обнаруживается вторичная структура. Конечно, если сделан рентген (или точный многомерный ЯМР) белка вторичная структура берется из атомных координат.
В течение более чем 25 лет основным методом получения расположения атомов, составляющих биомолекулу остается, метод РСА кристаллов и волокон биомолекул. Почему этот метод является столь информативным при определении структуры макромолекул? Обнаруженная Лауэ еще в начале ХХ века дифракция реентгеновских лучей позволяет определить положения атомов. Почему? Кристаллы являются (в принципе, любая более-менее упорядоченная структура) дифракционной решеткой по отношению к рентгеновским лучам. Рентгеновская дифракционная картина (рентгенограмма) является набором регулярно расположенных рефлексов (попросту - пятен). Эта картина - результат того, что рентгеновские лучи, проходя сквозь кристалл, отклоняются не в любых направлениях, а в строго определенных, зависящих от его строения, т. е. от параметров кристалла как дифракционной решетки. Если разрушить кристаллическую решетку, например, расплавить кристалл, регулярно расположенные дискретные рефлексы исчезнут - превратятся в диффузионные кольца (гало). Таким образом, рентгенограмма кристалла представляет собой зашифрованное изображение его регулярной, периодической структуры. Зная длину волны, можно по положениям рефлексов на рентгенограмме, т. е. по отклонению дифрагированных лучей, определить расстояние между периодически повторяющимися группами атомов в кристалле. Анализируя интенсивности этих рефлексов, можно определить и пространственное расположение всех атомов кристалла, от которого получена рентгенограмма. В начале 20-х годов ХХ века несколько групп исследователей, особенно - группа Астбюри, начали систематически изучать белки. Первые рентгенограммы были получены от фибриллярных белков - фиброина, кератина, миозина, коллагена. С этими белками было удобно работать, т. к. они существуют в виде довольно больших волокон или пластин, как фиброин, из которого построены нити натурального шелка, или кератина - белка волос, шерсти, рогов, ногтей. В 30-е годы на основе полученных рентгенограмм Астбюри были предложены два возможных типа структуры изученных белков: более компактный, или α- структура, и более растянутый, или β-структура.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
