

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Лекция 8. Вторичная структура белков
Каждый белок специфическим образом свернут в пространстве, и эта конформация определяет его физико-химические и биологические свойства. Пространственная структура белка в целом определяется его первичной структурой. Эта взаимосвязь создает предпосылки для теоретических расчетов и предсказаний вторичной структуры на основе их аминокислотной последовательности. Пространственная структура достаточно подвижна, т. е. способна изменяться под воздействием внешних условий или различных агентов. В этом смысле правильнее говорить о предпочтительной конформации белка, об одной из многих, энергетически более выгодной пространственной структуре.
Как мы уже говорили, главной структурной единицей белка или пептида является пептидная (амидная) связь - CO-NH-. Согласно современным представлениям она является практически плоской, ее основные параметры – на рисунке (см. презентацию Лекции). В обычных условиях наблюдаются лишь небольшие отклонения от плоскости системы (до 5-10о); большие деформации возможны в напряженных циклических системах. Пептидная связь на 10% короче обычной, простой C-N – связи и имеет характер “частично двойной” связи C=N-. Другими словами, в белках и пептидах связь C-N является частично кратной из-за взаимодействия неподеленной пары электронов атома азота с π-электронной системой карбонильной группы С=О, что приводит к затрудненному вращению вокруг C-N - связи (барьер вращения составляет 40 кДж/моль). Обычно пептидная связь имеет транс-конфигураци, т. е. является транспланарной (показать рис.). В напряженных циклических системах (производные пролина или при большом размере заместителей у атома азота) пептидная связь может существовать в плоской цис-конфигурации (показать рисунок).
Пептидная группа, точнее ее свойства, во многом определяют конформационные возможности полипептидной цепи. Геометрия полипептидной цепи определяется набором углов вращения. Три последовательные связи Сα-С, С-N и Сα-N имеют угла поворота ψ, ω и φ, соответственно. Из-за частично двойного характера связи С-N угол ω равен 180о. Поэтому конформационные возможности полипептидной цепи определяются набором значений углов ψ и φ. (Показать рисунки пептидной группы и основные углы вращения в ней). Т. е. знание значений углов ψ и φ для всех пептидных звеньев эквивалентно полной информации о пространственной структуре основной цепи белка. Графически конформационные параметры полипептидной цепи принято изображать с помощью карт, предложенных Рамачандраном еще в 1963г. и отражающих зависимость энергии аминокислотного остатка от параметров ψ и φ (рисунок показать). Значение углов ψ и φ откладывается по осям координат от –180о до +180о. В силу взаимодействия между боковыми цепями эти углы не могут принимать любые значения – для них разрешенными оказываются только некоторые дискретные области, которые соответствуют энергетически выгодным конформациям полипептидной цепи, т. е. являются областями минимума энергии.
Как мы уже говорили, аминокислотные цепи могут образовывать в пространстве высокоупорядоченные локальные структуры со спиральной симметрией. Возможность локализации спиральных участков по картам электронной плотности, полученными методом РСА, выделение таких участков с помощью различных физико-химических методов (т. к. такие участки стабилизированы с помощью ближних взаимодействий, например, Н-связей) позволяют изучить их независимо.
Рассмотрим сегодня вторичную структуру белков. Прежде всего у нас речь пойдет о регулярных вторичных структурах: об α-спиралях и о β-структуре, но не только о них. Укладка α и β-структур в глобулу определяет третичную структуру белка (Рис.7-1). Эти вторичные структуры отличаются определенными, периодическими конформациями главной цепи при разнообразии конформаций боковых групп.
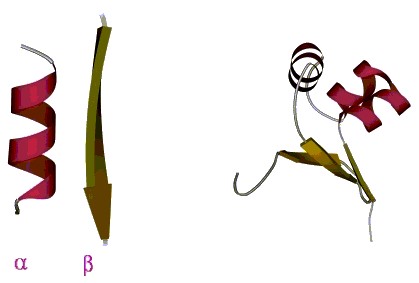
Рис.7-1. Вторичная структура полипептидной цепи (α-спираль и тяж β-листа) и третичная структура белковой глобулы.
Начнем со спиралей. Они могут быть левые и правые (Рис.7-2), у них может быть разный период и шаг. Правые (R) спирали приходят к нам, завиваясь против часовой стрелки (что отвечает положительному отсчету угла в тригонометрии); левые (L) - приходят, вращаясь по стрелке.
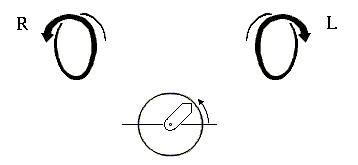
Рис.7-2. Правые (R) и левые (L) спирали. Под ними показан отсчет положительного угла в тригонометрии: при этом "близкая к нам" стрелка вращается против хода часов.
Важнейшие спирали в полипептидной цепи держатся водородными связями, где С=О группы остова полипептида связаны с лежащими от них в направлении С-конца цепи H-N группами. В принципе, возможны следующие спирали, стянутые Н-связями (Рис.7-3): 27, 310, 413 (обычно именуемая α) и 516 (она же π). Здесь в названии "27" - "2" означает связь со 2-м по цепи остатком (см. Рис.7-3), а "7" - число атомов в цикле (O......H-N-C'-Cα-N-C'), замыкаемом этой связью. Тот же смысл имеют цифры и в названии других спиралей.
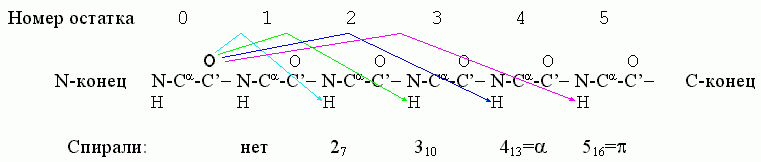
Рис.7-3. Водородные связи (они показаны стрелками), характерные для разных спиралей.
Наиболее известный элемент вторичной структуры - это α-спираль, впервые описанная Полингом и Кори в 1953 году. Она возникает под действием внутримолекулярных Н-связей между пептидными группами. Сворачивание полипептидной цепи в спираль обусловлено тремя цепочками водородных связей между пептидными группами. Первая цепь связывает Н первой пептидной группы с О 4-ой, водород 4-й - с кислородом 7-й и т. д. Вторая цепь Н-связей осуществляется между водородом 2-й пептидной группы и О 5-й, водородом 5-й и кислородом 8-й, Н 8-й и О 11-й и т. д. Третья цепь - Н 3-й с О 6-й, Н 6-й с О 9-й пептидной группы и т. д. В результате образуется правая спираль с шагом 5.6Е и диаметром 4.56 Е. Период спирали 27Е. На пять оборотов спирали приходится 18 аминокислотных остатков. Все боковые спирали расположены с наружной стороны спирали.
Интересно проследить за ходом рассуждений Полинга при попытке описать α-спираль. Длины связей и углы были известны из РСА-данных целого ряда малых пептидов. Разумно было предположить, что их значения не изменяются в любой вторичной структуре. Практически во всех известных структурах размеры пептидных групп одинаковы, а входящие в них имино - и карбоксильные группы находятся в плоской транс-конформации. Предположение, что такая конформация сохраняется всегда и во всех остатках, упростило дальнейшее рассмотрение и навело на мысль: если каждый пептидный скелет имеет одну и ту же структуру, то и весь полипептидный скелет должен обладать регулярной вторичной структурой типа какой-либо спирали. Чтобы ограничить количество возможных гипотетических моделей исследователи ввели такие ограничения:
1. количество Н-связей между пептидными группами N-H и C=O должно быть максимальным,
2. сами связи должны быть по возможности близки к линейным.
Какие из этих спиральных структур преобладают в белках? α-спирали. Почему? Ответ на этот вопрос дает карта Рамачандрана для типичного аминокислотного остатка аланина (Рис.7- 4), на которой отмечены конформации, периодическое повторение которых приводит к завязыванию изображенных на Рис.7-3 водородных связей.
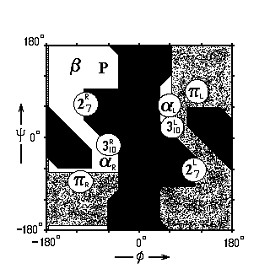
Рис.7-4. Конформации различных вторичных структур на фоне карты разрешенных и запрещенных конформаций аминокислотных остатков. 27R, 27L: правая и левая спираль 27; 310R, 310L: правая и левая спираль 310; αR, αL - правая и левая α-спираль; πR, πL - правая и левая π-спираль. β - β-структура (подробности см. на Рис.7-8б). Р - спираль Poly(Pro)II. - конформации, разрешенные для аланина (Ala); - области, разрешенные лишь для глицина, но не для аланина и других остатков; - области, запрещенные для всех остатков. φ и ψ - углы внутреннего вращения в белковой цепи.
Видно, что только спираль αR (α-правая) лежит достаточно глубоко внутри области, разрешенной для аланина (и для всех других остатков). Другие спирали лежат либо на краю этой области (например, левая спираль αL или правая спираль 310), где конформационные напряжения уже возрастают, либо в области, доступной только глицину.
Поэтому можно ожидать, что именно правая α-спираль должна быть, как правило, более стабильной, и потому преобладать в белках, что и наблюдается. В правой α-спирали (Рис.7-5) все атомы упакованы оптимально: плотно, но без напряжений; поэтому не удивительно, что в белках таких спиралей много, а в фибриллярных белках они достигают гигантской длины и включают сотни аминокислотных остатков.
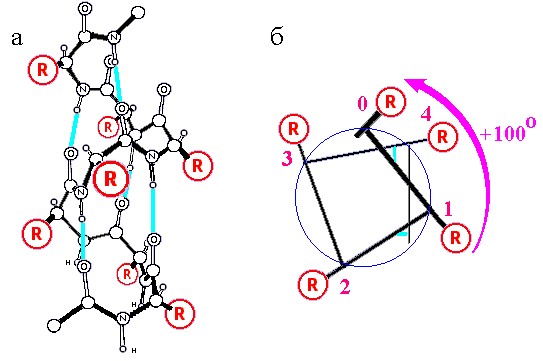
Рис.7-5. Правая α-спираль. (а) Атомарная структура. R - боковые группы. Голубые линии - водородные связи. (б) Схематическое изображение одного витка той же α-спирали, вид с торца. Стрелка показывает поворот спирали (в расчете на один остаток) по мере ее приближения к нам.
Левых α-спиралей в белках практически нет. Нет и спиралей 27, которые, мало того что лежат на самом краю разрешенной области, но еще имеют энергетически невыгодный, почти прямой угол схождения N-H и О=С групп.
Практически нет в белках и спиралей π. Они тоже лежат на самом краю разрешенной области, да еще и витки в них слишком широки, так что π-спирали имеют энергетически невыгодную пустую "дырку" на оси. А вот спирали 310 (в основном правые, левые пригодны практически лишь для глицинов) в белках есть, правда, в виде коротких (из трех-четырех остатков) и деформированных фрагментов (из-за стерических напряжений: слишком тугая спираль! - соответствующая ей конформация лежит на самом краю разрешенной области).
Отметим одно свойство спиралей, хорошо видное на Рис.7-5а: на их N-конце сидят свободные от внутриспиральных водородных связей Н атомы N-H групп, а на С-конце - свободные от водородных связей О атомы С=О групп. Так как электронное облако с Н атома частично стянуто электроотрицательным N атомом, а электроотрицательный О атом сам стягивает электрон с С' атома, N-конец спирали несет положительный, а С-конец - отрицательный парциальный заряд. То есть спираль представляет собой длинный диполь: величина суммарного (поставляемого тремя NH-группами) парциального "+" заряда на ее N-конце составляет около половины протонного, а "-" заряда на С-конце α-спирали около половины электронного заряда.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
