
Стойкость цГМФ в дифференцировке спутниковых клеток и эмбриональном миогенезе приводит к гипертрофии. Мы исследовали, является ли жестким регулирование NO / цГМФ сигнализации при дифференциации, чтобы предотвратить разрастание нефизиологических миофибрилл. Мы исследовали эту возможность, как в пробирке и в естественных условиях. Для имитации нерегулируемой цГМФ сигнализации в клетках-сателлитах, дифференциация была выполнена в постоянном присутствии 0.3-3 мм 8 Br-цГМФ. Кроме того, спутниковые клетки высевали при высокой плотности (3 ? 104 клеток/см2) для слияния. Через 48 ч дифференциации в этих условиях, спутниковые клетки имели признаки гипертрофической миотубы (рис. 3). Индуцированная гипертрофия зависит от концентрации, как это было с увеличением экспрессии миозина (рис. 3 C).Миотубы в 8-Br-цГМФ обработанных культурах считаются гипертрофированными, потому что их среднее число ядер и измерения размера волокна (рис. 3 б) и общее количество миозина (рис. 3 C) были значительно выше по сравнению с контрольной группой. Такая гипертрофия и увеличение миозина не наблюдалось в спутниковых клетках, дифференцировавшихся в присутствии 50-300 мкм DETA-нет, даже при более высоких концентрациях (рис. 3,-C, и не изображены).
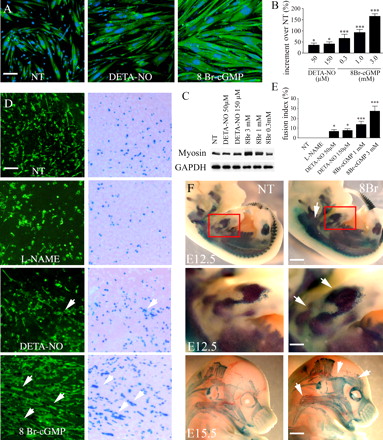
Дифференцировкиа спутниковых клеток происходит через сигнальные пути схожие, но не идентичны тем, которые наблюдались во время эмбрионального развития скелетных мышц (Заряд и Рудницкий, 2004). Поэтому мы оценили роль NO-цГМФ пути и его дерегулирования в эмбриональном и фетальном миогенезе. Пресомитная мезодерма (PSM) или хвостовые сомиты (I-III) (MLC1/3F)-nLacZ 9,5-D эмбрионов трансгенных мышей, в которых ген LacZ находится под контролем транскрипционных факторов MLC1/3F (Kelly и соавт., 1995), были выращены в качестве эксплантов в культуру в присутствии или в отсутствии 50-300 мкм DETA-NO, 1-3 мм 8 Br-цГМФ, или 5 мм L-NAME. После введения всего лишь 2 г в культурах, дифференцированные мононуклеарные миоциты полученны из PSM эксплантов, и отмечено увеличение числа клеток
![]()
после дополнительных 2 г (рис. 3, D и E). Ни 50-300 мкм DETA-NO, ни L-NAME не давали соответствующего воздействия на время появления и количество миоцитов после 4 дней испытания при любой концентрации (рис. 3, D и E, а также не показан), в отличие от 3 мм 8 Br-цГМФ, что вызвало формирование миотуб, события, которые не происходят в физиологических PSM эксплантах культур (Cossu и Biressi, 2005), что свидетельствует об ограничении слияния при помощи циклических нуклеотидов (рис. 3 D). Эффект от 8 Br-цГМФ зависит от концентрации (рис. 3 E). Сходные результаты были получены в сомитах эксплантов (неопубликованные данные).Для изучения миогенеза в конце развития эмбриона и плода, MLC1/3F-nLacZ беременных мышей лечили с или без 8 Br-цГМФ (3 г / кг массы тела) на 10-й день беременности либо 12,5 или 15,5. Затем миогенной клетки эмбрионов были выявлены путем окрашивания LacZ. Как показано на рис. 3 F, 8 Br-цГМФ-обработанные эмбрионы показали расширенное окрашивания LacZ, указывая на повышенный уровень миогенеза в обеих временных точках. Эти результаты ясно показывают, что постоянное присутствие цГМФ увеличивается размер и приводит к гипертрофии мышц и о том, что жесткое регулирование его концентрации необходимо для нормального процесса.
NO / цГМФ сигнализация в миогенезе опосредовано регулирует экспрессию фоллистатина через активацию транскрипции опосредовано NFAT / CREB / MyoD. Мы были заинтересованы в выявлении молекулярных эффекторов NO / цГМФ сигнализации и оценке насколько молекулы , такие как IGF-I, интерлейкина-4 (ИЛ-4;. Хорсли и др., 2003), или follistatin (. Iezzi и др., 2004), играют важную роль в мышечной гипертрофии. Таким образом, выполняется полуколичественный ОТ-ПЦР на РНК, выделенной из дифференцированных клеток спутника, PSM эксплантов, или мышц на эмбриональном и фетальном этапах, которые были культивированы с 3 мм 8 Br-цГМФ, 50 мкМ DETA-NO, или 5 мм L-NAME использованием праймеров, специфичных для IGF-I, IL-4, фоллистатина, миостатина, и скелетных мышц MLC1/3F (рис. 4). Параллельно мы исследовали экспрессию IGF-I, IL-4, фоллистатина, миостатина, и тяжелой цепи миозина методом вестерн-блоттинга (рис. 4 B). Количественная оценка и статистический анализ полученных результатов показан на рис. S1 (доступно на http://www. jcb. org/cgi/content/full/jcb.200507083/DC1).
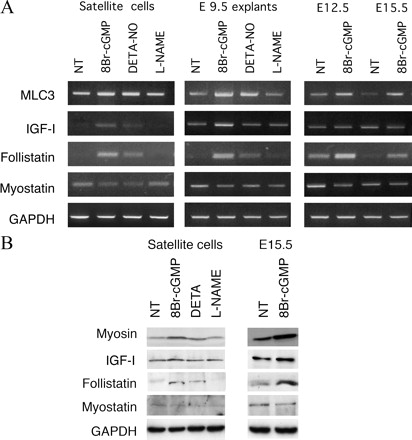
Хотя L-NAME не имел эффекта в любых условиях, мы наблюдали повышение уровня транскриптов легких цепей миозина вах 8 Br-цГМФ-и DETA-NO-обработанных культур и эмбрионах. В сателлитных клетках это сопровождалось заметным увеличением в экспрессии
![]()
фоллистатина. Важное значение в PSM эксплантах и эмбрионах - экспрессия фоллистатина увеличилась только в присутствии 8 Br-цГМФ, в то время как DETA-NO не оказали существенного влияния (рис. 4 и рис. S1). Уровни белка миостатина не оказывают существенного влияния, даже если бы было снижение уровня мРНК. Мы также отметили, что мРНК и уровни IGF-I были немного увеличены при обработке 8 Br-цГМФ. Эти изменения, однако, не были значимыми (рис. 4 и рис. S1). Никаких изменений в уровнях IL-4 не было обнаруженыо(неопубликованные данные). Эти результаты позволяют предположить, что увеличение фоллистатина имеет отношение к NO / цГМФ сигнализации во время дифференциации миобластов. Фоллистатин недавно был описан как центрального медиатора фузогенного эффекта, который оказывает ингибитор дезацетилазы на слияние миобластов в миотубы через пути отличны от тех, которыми действует или IGF-I или IL-4 (Iezzi и соавт., 2004). В частности, регулирование ингибиторами дезацетилазы фоллистатина, как представляется, совместно активирует MyoD, ядерный фактор активированных Т-клеток (NFAT) и цАМФ связывающий белок (CREB;. Iezzi и др., 2004).Для исследования того, может ли NO-цГМФ путь активации промоутера фоллистатина действовать тем же путем, мы использовали миогенную клеточную линию C2C12. Как и сателлитные клетки, миобласты C2C12 привели к образованию гипертрофическоих миотуб при культивировании в постоянном присутствии 8 Br-цГМФ (рис. 5, А и B). Это сопровождалось зависимостью от концентрации увеличения уровня фоллистатина (рис. 5 C). Для изучения влияния на транскрипцию фоллистатина, промоутер связанный с геном люциферазы (FS-Luc) трансфицировали в C2C12 клетки (Iezzi и соавт., 2004). 8 Br-цГМФ активировал транскрипцию Fs-Luc (рис. 5 D). Чтобы установить, является ли активация промотора фоллистатина цГМФ опосредованной MyoD, CREB, и NFAT, FS-Luc-трансфицированные C2C12 были дифференцированы в присутствии 8 Br-цГМФ и / или трансфицированные негативным регулятором MyoD; Id1 (Iezzi и др., 2004);.. или доминантно-негативной формой CREB, A-CREB (Herzig и др., 2001), или Vivit, который представляет собой пептид, блокирующий NFAT-зависимоую транскрипцию (Aramburu и др., 1999).. Мы заметили, что 8-Br-цГМФ-зависимой активации Fs-Luc подавляется в присутствии любого из этих ингибиторов (рис. 5 D). Таким образом, NFAT, MyoD, и CREB оказывают опосредованные эффекты 8 Br-цГМФ на транскрипцию фоллистатина. Предыдущие исследования других типов клеток показали, что CREB и NFAT активируются через цГМФ, протеинкиназу G-зависимого фосфорилирования (Гуди и др., 1996;. Pilz и Casteel, 2003; Гонсалес Боск и др., 2004.). Соответственно, мы обнаружили, что 3 мкМ каждого из двух ингибиторов структурно связаных протеинкиназ G, KT5823 и 8-( chlorophenylthio) гуанозин 3 ', 5'-циклического монофосфоротиоат ([Rp]-8-pCPT-cGMPS;. Смоленский и др., 1998 ), позволили индукцию FS-Luc (рис. 5 D). В гладких мышцах цГМФ может действовать через протеинкиназу А, фермента, который играет важную роль в выбранном миогенном пути (Корнуэлл и др., 1994;.. Chen и др., 2005). Возможность участия протеинкиназы в посредничестких эффектах цГМФ, однако, была исключена в связи с отсутствием ингибирующих эффектов при 3 мкМ ингибиторов KT5720 и (Rp)-8-pCPT (рис. 5 D) .
![]()
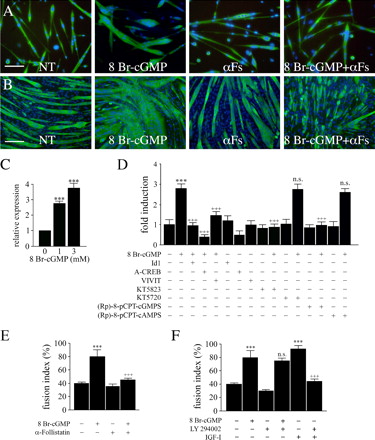
Затем мы изучали может ли гипертрофический эффект 8 Br-цГМФ быть исключительно зависит от активации транскрипции фоллистатина, IGF-I был также вовлечен в силу небольших, но обнаруживаемых изменений в уровне IGF-I транскриптов индуцированных при воздействии 8 Br-цГМФ (рис. 4). Мы культивировали как спутниковые, так и C2C12 клетки в присутствии 8 Br-цГМФ и либо антител, направленных против фоллистатина, чтобы нейтрализовать его (Iezzi и соавт., 2004) или фосфатидилинозитол-3 "ингибитора киназы LY294002, который ингибирует IGF - 1 сигнализацию в мышцах (Ибарра и соавт., 2004). Нейтрализующие антитела к фоллистатину ингибируют действие 8 Br-цГМФ без значительного воздействия на спутниковые клетуи (рис. 5, А и Е) и C2C12 клетки (рис. 5 б). Эти результаты показывают, что эффект цГМФ в основном опосредован активацией фоллистатина. В соответствии с этим, LY294002 не изменяет эффекты 8 Br-цГМФ в то время как ингибирование IGF-I в спутниковых клетках и C2C12 (рис. 5 F). Для оценки эффекта клеточной специфичности цГМФ на фоллистатин, мы исследовали, может ли он также увеличивать экспрессию фоллистатина в клетках немышечных линий, а именно эмбриональных клеток карциномы (P19), взрослых (NIH 3T3) и эмбриональных (10 T1 / 2) фибробластов, мезангиобластов, полученных предшественников гладких мышц (D351;. Brunelli и др., 2004), а также первичных культур кардиомиоцитов. 8 Br-цГМФ не имел влияния на уровни фоллистатина в любой из этих клеток, в то время как постоянно возрастающая индукция фоллистатина в C2C12 клетках использовали в качестве контроля (рис. S3, http://www. jcb. org/cgi/content/full / jcb.200507083/DC1).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
