
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

Индукция фоллистатина при помощи оксида азота через цГМФ сигнальный путь, который контролирует слияние миобластов.
Скелетная мышечная ткань состоит из многоядерных волокон, которые возникают в определенные периоды эмбриогенеза от слияния миобластов (Buckingham соавт., 2003). В частности, эмбриональных и фетальных миобластов, происходящих из различных популяций мио-бластов (Cusella-De Angelis и др., 1994;.. Relaix и др., 2005), приводят к появлению первичных(примерно [E] 11-12 эмбрионального развития) и вторичных (около E15-16) волокон, соответственно (Ontell и Kozeka, 1984). Впоследствии, мышечная масса подвергается экстенсивному росту в пренатальный и послеродовой период, и этот рост поддерживают специализированные клетки, клетки-сателлиты, расположенные в нише между плазмалеммой и базальной пластинкой волокон (Таджбахш, 2003). На протяжении миогенеза баланс между пролиферацией, дифференцировкой и слиянием необходим для правильного формирования окончательных структур мышцы (Тацуми и др., 2002;.. Букингемского и др., 2003). Многие положительные и отрицательные сигналы, ответственные за регулирование такого тонкого баланса, действующие на эмбриональном/ пренатальном этапе и после родов. Они включают в себя факторы транскрипции, такие как MyoD, Myf5, MRF4, и миогенин, а также внеклеточные агонисты и антагонисты, такие как члены семейства инсулиноподобного фактора роста IGF и TGF, FGF, фактора роста гепатоцитов, и костного морфогенетического белка (BMP) и его антагонистов (Balemans и Ван Хул, 2002;. Parker и др., 2003). Оксид азота (NO) регулирует основные функции взрослых скелетных мышц, такие, как деятельность нервно-мышечного синапса, связь возбуждение-сокращение, расширение кровеносных сосудов, глюкозы, функции митохондрий и биогенез, гликолиз (Wang и др. ., 1995; Balon и Надлер, 1997; Клементи и Meldolesi, 1997; Wolosker и др., 1997;. Bredt, 1998; Stamler и Мейснер, 2001; Eu и др., 2003;. Nisoli и др., 2004). О возможности того, что NO играет важную роль в скелетном миогенезе можно судить по наблюдениям, что он участвует в активации спутниковых клеток (Anderson, 2000;. Тацуми и др., 2002), а его синтез фермента NO-синтазы (СОС), которая регулирует развитие и может привести к активации миогенной программе IGF-II (Lee и др., 1994;. Blottner и удачи, 1998; El Dwairi и др., 1998;.. Калиман и др., 1999). Точная роль NO в миогенезе и сигнальных путях, однако, не известно. В настоящей работе мы исследовали эти аспекты, как в пробирке и в естественных условиях, на разных этапах миогенеза. Наши результаты показывают, что NO непосредственно стимулирует слияние миобластов до регулирования фоллистатином. Мы также обнаружили, что действие NO ограничено определенным временным окном и опосредовано через жесткую регуляцию активации гуанилатциклазы и генерации циклического гуанозинмонофосфата (цГМФ) (Монкада и соавт., 1991). цГМФ сигнализация при лечении 8 Br-цГМФ приводит к увеличению процессов слияния с образованием гипертрофических миотуб и мышечных волокон в пробирке и в естественных условиях. В целом, наши результаты свидетельствуют о ключевой роли NO / цГМФ в регуляции слияния миобластов во время мышечного развития.
Результаты
Экзогенная и эндогенная NO увеличивает спутниковую активацию в пробирке в определенном временном окне. Для изучения влияния NO на дифференциацию и слияние миобластов, спутниковые клетки, выделенные у новорожденных мышей и нанесенные низкой плотностью на пластины (6 ? 103 клеток/см2), поддерживали в течение 48 ч в питательной среде, а затем перенесли в среду дифференциации в присутствии или отсутствии повышенной концентрации донора NO (Z) -1 - [2 - (2-аминоэтил)-N-(2-ammonioethyl) амино] diazen-1-ий-1 ,2-diolate] (DETA-NO), , который высвобождает NO постоянно и в определенных концентрациях (Клементи и соавт., 1998), или N?-нитро-L-аргинин метилового эфира (L-NAME), который является ингибитором широкого спектра NOS (Монкада и соавт., 1991). Индекс слияния измеряют через 24, 48 или 72 часов. Как показано на рис. 1 A, DETA-NO увеличилась, в то время как L-NAME снизилась, индекс слияния измерялся способами в зависимости от концентрации. Эти эффекты были конкретными, поскольку соответствующий амин, DETA, не дают значительного эффекта и действия L-NAME ингибируется 5 мМ L-аргинином (неопубликованные данные). Стимуляция DETA-NO и ингибирование L-NAME были четко обнаружены через 24 ч и увеличили дифференциацию, которая становится статистически значимой после 48 и 72 ч инкубирования культуры (рис. 1, B и C). Через 72 часа почти нет событий слияния среди миозин-экспрессирующих клеток-сателлитов в присутствии 5 мМ L-NAME (Рис. 1 C). Тот факт, что последствия DETA-NO и L-NAME влияния на слияние присутствовали уже после 24 ч лечения, предполагает, что эти события имели место на ранней стадии дифференциации. Для установления временного окна, 5 мМ L-NAME и 50 мкМ DETA-NO (уступая концентрации 120 ± 5 нм, N = 5, измеренная с NO электродом;. Клементи и др., 1998) были добавлены к дифференцирующимся клеткам сателлиттам в различные моменты времени. Индекс слияния оценивали после 72 часов. Как показано на рис. 1 D, как DETA-NO и L-NAME были максимально эффективны в укреплении и предотвращения слияния, соответственно, при добавлении в начале процесса дифференцировки. Соединения были все менее эффективными при добавлении в более поздние моменты времени и почти полностью неэффективными, когда добавлялись после 16 часов. Последовательно мы обнаружили, что процесс дифференциации сопровождается ранним повышением активности NOS, пик которого пришелся на 8 часов, а потом снижалсь, вернувшись в базальным уровням после 48 ч (рис. 1 E). Мы обнаружили, что клетки-сателлиты экспрессируют эндотелиальную (NOS III) и мышечную (NOS I?) вариантов нейронных NOS. Уровни экспрессии NOS III и I? были неизменными на протяжении процесса дифференцировки (рис. 1 F), указывая, что изменения в активности NOS были последствиями активации и ингибированием активности фермента, а не изменения в экспрессии белка. NOS II экспрессия не была обнаружена на протяжении всего времени анализа (неопубликованные данные).
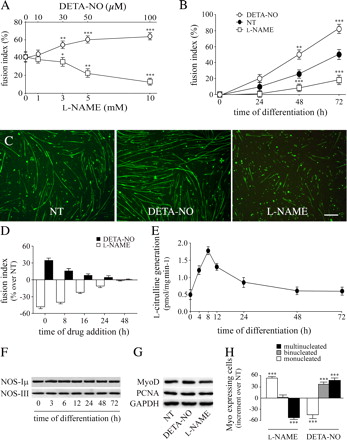
Спутниковые клетки культивировали в пробирке асинхронно, небольшая часть из них все еще сохранялась, как недифференцированные клетки даже после нескольких дней в
![]()
дифференцирующей среде (Cossu и др., 1980;. Ontell и Kozeka, 1984). Увеличение слияния клеток-сателлитов вызваное NO может быть связано с влиянием этого посланника на сам процесс или вторичным по отношению к NO-зависимым недифференцированным клеткам в стадии терминальной дифференцировки (Anderson, 2000). Это должно привести к увеличению MyoD экспрессии и соответствующему снижению клеточной пролиферации. Чтобы различать эти возможности, мы изучили влияние NO на эти процессы. Как показано на рис. 1 G, воздействие на спутниковые клеточные культуры в течение 24 часов либо 5 мМ L-NAME или 50 мкм DETA-NO не изменило ни экспрессии MyoD маркером, ни ДНК-полимеразы ? кофактора (PCNA), который экспрессируется в S фазе клеточного цикла. Мы не обнаружили ни увеличения, ни распространения, измеренные путем подсчета числа ядер во всех клетках, или числа ядер, присутствующих в миозин-позитивных клетках (неопубликованные данные). Это исключает возможность того, что образование миотубы с увеличенным количеством ядер и с увеличенным размером явилось следствием увеличения числа недифференцированных клеток в стадии терминальной дифференцировки, предполагая, что не было никаких актов непосредственно в качестве индуктора слияния миобластов. Процесс, при котором клетки переходят в мышечные волокна, является следствием начального слияния между двумя миобластами и последующим слиянием новых клеток (Заряд и Рудницкий, 2004). Как показано на рис. 1 Н, 5 мМ L-NAME увеличило, в то время как 50 мкМ DETA-NO уменьшило долю мононуклеарных клеток. Кроме того, DETA-NO увеличил образование двуядерных клеток и многоядерных мышечных трубок, в то время как L-NAME сократило число миотуб. Результаты, представленные на рис.1 показали, что активность NOS и генерация NO которые жестко регулировались во время спутниковой дифференцировки клеток и не вызывали и не усиливали слияние без влияния на другие события в программе дифференциации этих клеток. Тот факт, что экзогенный NO не является эффективным, когда NOS блокирована, предпологает, что NO сигнализация регулируется не только на уровне своего уровня NOS, но и ниже его.
Влияние NO на слияние спутниковых клеток зависит от цГМФ. Далее мы проанализировали зависимость эффектов NO на активацию гуанилатциклазы и генерацию цГМФ(Монкада и соавт., 1991). Как показано на рис. 2 A, дифференциации клеток-сателлитов в культуре сопровождается генерацией цГМФ, происходящей в 4-24-ч, в соответствии с временным окном влияния NO (рис. 1 E). Большое значение придается способности DETA-NO увеличивать концентрацию циклических нуклеотидов, что значительно выше при введении в течение первых 12 часов процесса дифференцировки (рис. 2, сравнить время 72 ч со временем 0). Эти результаты позволяют предположить, что в сателлитных клетках чувствительность гуанилатциклазы к NO регулируется и ее активации происходит на начальных этапах дифференцировки. Поскольку уровни экспрессии гуанилатциклазы подвидов ? и ? не меняется со временем (рис. 2 Б), кажется, что такое регулирование происходит через каскад посттрансляционных событий
![]()
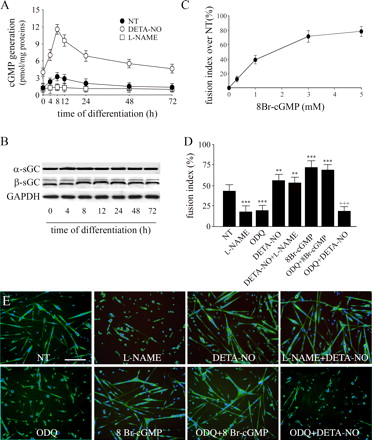
Для оценки роли цГМФ в области слияния спутниковых клеток мы изучали влияние клеточной мембраны, 8 Br-цГМФ (0,3-5 мм) и ингибитора гуанилат циклазы 1Н-(1,2,4) оксадиазоло [4,3-?] хиноксалин-1-он (ODQ, 3 мкм), которые были введены в временное окно 4-24-ч, в котором эти клетки генерируют цГМФ. Слияние спутниковых клеток измеряли после 72 часов. Как показано на рис. 2 (C-E), 8 BrcGMP и ODQ имитировали эффект DETA-NO и L-NAME, соответственно. Эти результаты ясно показывают, что эффект NO на слияние спутниковых клеток зависит от активации гуанилатциклазы и генерации цГМФ. Результат изображен на рис. 1, свидетельствует об отсутствии сигнализации не только на уровне своего поколения NOS, но и далее по ходу каскада реакций от нее. Результаты на рис. 2, показывающие зависимость эффекта NO на цГМФ и зависящие от времени изменения в чувствительности гуанилатциклазы, показывают, что основным в этом каскаде является цГМФ.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
