
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Деятельности дыхательного центра свойственна высокая степень надежности. В ее обеспечении участвуют афферентные импульсы, способствующие смене дыхательных фаз.
Сокращения многих мышц и мышечных групп, участвующих в вентиляции легких, строго координированы по протеканию во времени и силе. Интенсивность возбуждения дыхательных мышц регулируется в соответствии с изменениями их длины и объема грудной клетки. Эти стороны деятельности аппарата внешнего дыхания обслуживаются рефлексами, рецептивные поля которых расположены в самом дыхательном аппарате: в легких, дыхательных мышцах, верхних дыхательных путях. Они выполняют функцию обратной связи между центрами и периферией и должны быть отнесены к собственным дыхательным рефлексам, осуществляющим саморегуляцию дыхания.
Рассмотрим строение рефлекторных дуг этих рефлексов.
Афферентная система легких. В 1868 г. Геринг и Брейер обнаружили, что увеличение объема легких тормозит сокращение мышц вдоха, а отсасывание воздуха из легких, наоборот, вызывает сильное сокращение инспираторных мышц. Зависимость деятельности дыхательного центра от объема легких устраняется двусторонней перерезкой блуждающих нервов или только их легочных ветвей.
Имеется несколько видов механорецепторов в легких. Морфологи различают медленно и быстро адаптирующиеся рецепторы растяжения легких, рецепторы спадения легких, рецепторы слизистой оболочки трахеи и бронхов, рецепторы интерстициальной ткани альвеол (т. н. Ю-рецепторы легких). Роль и значение всех этих образований в регуляции дыхательных движений различна.
Изменения объема легких у животных вызывает три сильные и постоянные реакции дыхательного центра: 1) торможение инспираторной активности при увеличении объема легких, 2) короткое инспираторное возбуждение при резком и небольшом увеличении объема и 3) увеличение частоты дыхания и силы сокращений мышц вдоха при уменьшении объема легких. Для этих рефлексов характерны системные реакции дыхательного аппарата, причем состояние мотонейронов мышц вдоха и выдоха изменяется реципрокно.
Хотя двусторонняя ваготомия не приводит к смерти животного. но выключение импульсов от рецепторов легких существенно меняет протекание дыхательных периодов и форму дыхательных движений. Происходит увеличение амплитуды и продолжительности вдохов и выдохов, а смена дыхательных фаз нарушается и происходит за счет раздражения механорецепторов дыхательных мышц. Афферентная система легких играет важную роль в саморегуляции дыхания. являясь основой обратной связи между периферией дыхательного аппарата и центрами.
Афферентная система дыхательных мышц. Диафрагма относительно бедна рецепторами, которые в обычных условиях не имеют существенного значения в регуляции дыхания. Зато дыхательная активность диафрагмы находится в постоянной зависимости от объема легких. При герметичной плевральной полости движения диафрагмы всегда сопровождаются раздражением механорецепторов легких, которые, по существу. заменяют собственные рецепторы диафрагмы.
Межреберные мышцы снабжены большим количеством рецепторов типа мышечных веретен. В мышцах одного межреберного промежутка насчитывают до 100 таких образований. Возбуждение окончаний веретен изменяется при сокращении и растяжении межреберных мышц. От чувствительных окончаний веретен в спинной мозг постоянно поступает поток импульсов, который усиливается при вдохе, так как вместе с экстрафузальными мышечными волокнами при вдохе происходит сокращение и интрафузальных, причем начало сокращения последних определяется раньше, чем возбуждение альфа-мотонейронов. Активность мотонейронов мышц вдоха и выдоха изменяется строго реципрокно.
Кроме рецепторов растяжения мышц, при дыхательных движениях происходит раздражение механорецепторов кожи грудной клетки, и рецепторов подкожных вен. Импульсы от механорецепторов грудной клетки поступают в грудные сегменты спинного мозга, восходят к диафрагмальным центрам и в головной мозг.
Нормальный дыхательный объем обеспечивается укорочением дыхательных мышц, развивающих определенное напряжение. Дыхательный центр определяет "запрос" на укорочение дыхательных мышц через эфферентные системы мышечных веретен. Сокращение интрафузальных волокон обуславливает дополнительное сокращение экстрафузальных мышечных волокон, пропорциональное укорочению интрафузальных волокон в соответствии с запросом. При увеличении нагрузки дыхательного аппарата (увеличение сопротивления дыханию) прежнее напряжение мышц не обуславливает прежнего укорочения и необходимого изменения объема грудной полости. Но в этих условиях веретена оказываются более растянутыми, чем до нагрузки, что в порядке рефлекса растяжения автоматически вызывает увеличение напряжения мышц.
Хеморецепторы дыхательного аппарата. Помимо механорецепторов легких и воздухоносных путей, а также проприорецепторов дыхательных мышц большую роль в регуляции дыхания играют сенсорные образования, чувствительные к химическим раздражителям, хеморецепторы. Функция последних - контроль газового состава и кислотно-щелочного баланса внутренней среды организма, в обеспечении постоянства которой дыхание принимает прямое участие.
Интенсивность внешнего дыхания в конечном счете определяется динамикой потребления кислорода и продукцией СО2 тканями тела. Дыхательный центр продолговатого мозга поддерживает уровень легочной вентиляции прежде всего в соответствии с напряжение углекислого газа и концентрацией водородных ионов в омывающей его крови. Однако этот центр, если его изолировать от афферентных связей с периферией, не способен адекватно реагировать на уровень доставки кислорода. Именно хеморецепторы посылают в дыхательный центр сигналы о величине напряжения кислорода в крови, а также дополнительную информацию о напряжении углекислоты и активной реакции внутренней среды. Показано, что эти рецепторы чувствительны к ограничению кислородного снабжения и снижению содержания кислорода в крови независимо от того, каким путем оно происходит.
Рецепторы, воспринимающие газовый состав артериальной крови, расположены в двух областях: дуге аорты и в каротидном синусе (место деления сонной артерии на наружную и внутреннюю). Хеморецепторы заключены в особых телах - клубочках, или гломусах, которые находятся вне сосуда и омываются кровью через специальные капилляры.
Кроме этих рецепторов, в регуляции газового состава крови принимают участие т. н. центральные нейрорецепторные образования. Перфузия 4 мозгового желудочка животных подкисленными или насыщенными СО2 растворами вызывает гипервентиляцию. Исследования показали, что хемочувствительные области располагаются в вентролатеральной части продолговатого мозга, на глубине 2,5-3 мм от поверхности, и посылают информацию нейронам дыхательного центра.
Благодаря функциональным свойствам артериальных хеморецепторов стимуляция их особенно эффективна при мышечной деятельности, которая, как известно, требует поддержания высокого уровня вентиляции. При этом хеморецепторы участвуют в регуляции не только МОД, но и таких параметров, как тонус бронхиальной мускулатуры и просвет воздухоносных путей, а также - путем влияния на активность межреберных мышц - на функциональную остаточную емкость и структуру дыхательного цикла.
Аортальные хеморецепторы расположены у "ворот" всей артериальной системы, а каротидные - у "ворот" сосудистой сети головного мозга. Исключительная важность функции каротидного тела указывает на большую физиологическую значимость регуляции газового состава крови, снабжающей мозг.
Дыхательный центр ретикулярной формации мозгового ствола осуществляет интеграцию поступающих хеморецепторных сигналов с другими афферентными и центральными влияниями. Полагают, что в результате взаимодействия механорецепторных и хеморецепторных импульсов в специализированных нейронных сетях и формируется специфический ритмический характер деятельности дыхательного центра.
Как же устроен дыхательный центр, который осуществляет столь тонкую регуляцию дыхания организма? Мы уже несколько раз упоминали о нем, давайте теперь поговорим более подробно.
Дыхательным центром называют совокупность нервных клеток, расположенных в разных отделах ЦНС, обеспечивающих координированную ритмическую деятельность дыхательных мышц и приспособление дыхания к изменяющимся условиям внешней и внутренней среды организма. Некоторые группы нервных клеток являются совершенно необходимыми для ритмической деятельности дыхательных мышц. Они расположены в ретикулярной формации продолговатого мозга, составляя дыхательный центр в узком (анатомическом) смысле слова. Нарушение функции этих клеток приводит к прекращению дыхания вследствие паралича дыхательных мышц.
Анализируя результаты перерезок, электрического раздражения и коагуляции различных участков продолговатого мозга, Миславский (1885) пришел к заключению, что дыхательный центр (ДЦ) находится в ретикулярной формации продолговатого мозга по обеим сторонам шва на уровне корней подъязычного нерва. Клеточные структуры центра простираются от нижнего угла почли до основания писчего пера. С боков они ограничены веревчатыми телами, а снизу оливами и пирамидами. Миславский доказал, что дыхательный центр имеет инспираторную и экспираторную части (центр вдоха и центр выдоха). В настоящее время показано, что инспираторные нейроны преобладают каудальном отделе tractus solitarius, экспираторные - в вентральном ядре (nucleus ambiguus).
Дыхательный центр имеет достаточно сложную организацию. Клеточные элементы, принимающие участие в интеграции дыхательного акта, представляют собой две функционально неоднородные группы нейронов, составляющие воспринимающий исполнительный отдел дыхат5льного центра. Воспринимающие структуры состоят из цепей инспираторных и экспираторных нейронов, имеющих двусторонние связи как между собой, так и с функционально однотипными нейронами противоположной стороны. Сигнализация с рецепторов дыхательного аппарата и с сосудистых рефлексогенных зон, а также из высших отделов мозга, воспринимаются главным образом этими нейронами.
Исполнительный отдел дыхательного центра также является парным образованием. Он представлен двумя группам нейронов, одна из которых более тесно связана с функцией диафрагм, другая – с функцией межреберных дыхательных мышц. Эти связи осуществляются через системы вставочных нейронов, с которыми контактируют нейронные пути, идущие от коры головного мозга, мозжечка и других центральных структур.
Лумсден и другие исследователи в опытах на теплокровных животных показали, что ДЦ имеет более сложную структуру, чем предполагалось ранее. В верхней части варолиевого моста они обнаружили т. н. пневтомотаксический центр который контролирует деятельность расположенных ниже в продолговатом мозге центров вдоха и выдоха. Между инспираторными и экспираторными нейронами существуют реципрокные отношения. Это значит, что возбуждение одной группы нейронов тормозит деятельность другой и наоборот.
Взаимодействие между нейронами ДЦ в настоящее время представляется следующим образом. Вследствие рефлекторных импульсом с хеморецепторов возникает возбуждение инспираторных нейронов и реципрокное торможение экспираторных. Одновременно импульсы от инспираторных нейронов поступают к центру пневмотаксиса, а от него к экспираторным нейронам, вызывая их возбуждение и акт выдоха. Одновременно центр выдоха возбуждается импульсацией с рецепторов растяжения легких. Активация экспираторных нейронов реципрокно тормозит инспираторный центр, но через центр пневмотаксиса наступает новое его возбуждение, подкрепляемое импульсацией от рецепторов спадения легких.
Деятельность всей совокупности нейронов, образующих ДЦ, необходима для сохранения нормального дыхания. Однако в процессах регуляции дыхания принимают участие также вышележащие отделы ЦНС, которые обеспечивают тонкие приспособительные изменения дыхания при различных видах деятельности. Важная роль в регуляции дыхания принадлежит большим полушариям головного мозга и их коре, благодаря которой осуществляется приспособление дыхательных движений при разговоре, пении, спорте и трудовой деятельности. Способность коры головного мозга влиять на процессы внешнего дыхания видна из того, что можно произвольно менять частоту и ритм дыхания, и, кроме того, можно выработать условно-рефлекторные изменения дыхания (например, предстартовые изменения дыхания у спортсменов и т. п.).
Зависимость деятельности дыхательного центра от газового состава крови. Деятельность ДЦ, определяющая частоту и глубину дыхания, зависит прежде всего от напряжения газов. растворенных в крови, и концентрации в ней водородных ионов. Ведущее значение в определении величины вентиляции легких имеет напряжение СО2 в артериальной крови. Для обозначения повышенного, нормального и сниженного напряжения СО2 в крови используют термины «гиперкапния», «нормокапния» и «гипокапния» соответственно. Нормальное содержание кислорода называется нормоксией, а недостаток кислорода в организме и тканях – гипоксией. Состояние, при котором гиперкапния и гипоксия существуют одновременно, называется асфиксией. Гипокапния и повышение уровня рН крови ведут к уменьшению вентиляции, а затем и остановке дыхания – апноэ.
 |
Кора головного мозга
Гипоталамус
Бульбарные центры
 |
Рисунок. 15. Функциональная система кислородного обеспечения организма (ФСКС)
Развитие гипоксии вначале вызывает умеренное увеличение вентиляции, а основном за счет возрастания частоты дыхания. Причиной такой реакции является рефлекторное возбуждение углекислотой хеморецепторов каротидного синуса и хемочувствительных клеток самого дыхательного центра. При увеличении степени гипоксии гиперпноэ сменяется ослаблением дыхания и его остановкой. Апноэ вследствие гипоксии смертельно опасно. Его причиной является ослабление окислительных процессов в мозге, в том числе и в нейронах дыхательного центра. Гипоксическому апноэ предшествует потеря сознания.
В целом поддержание нормального газового состава крови основано на принципе отрицательной обратной связи. Так, гиперкапния вызывает усиление активности дыхательного центра и увеличение вентиляции легких, а гипокапния – ослабление деятельности дыхательного центра и уменьшение вентиляции.
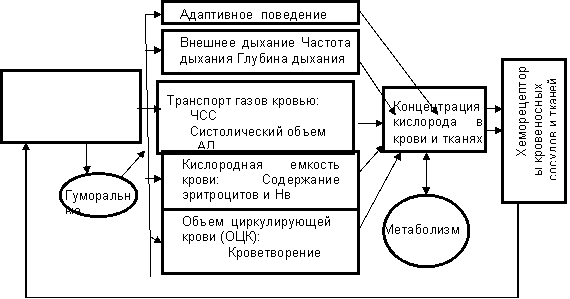
Функциональная система кислородного снабжения организма.
До сих пор мы рассматривали лишь регуляцию внешнего дыхания. Однако для поддержания нормального уровня концентрации кислорода в крови одного внешнего дыхания недостаточно. В число исполнительных механизмов функциональной системы кислородного снабжения организма (ФСКС) входят еще механизмы, обеспечивающие связывание кислорода, его транспортировку, уровень окислительно восстановительных процессов, а также серию поведенческих проявлений, направленных на сохранение кислородного снабжения. Естественно, что системообразующим фактором в ФСКС выступает уровень кислорода в крови, который контролируется хеморецепторами. Схема ФСКС представлена на рисунке 15.
Наиболее наглядно вовлечение различных исполнительных механизмов ФСКС в реализацию полезного результата - обеспечения нормального содержания кислорода в крови - проявляется при различных экстремальных условиях, к которым прежде всего относятся условия пониженного или повышенного атмосферного давления.
Функциональная система питания. Регуляция пищеварительных процессов
Пищеварением называется совокупность физиологических, физических и химических процессов, обеспечивающих прием и переработку поступающих из внешней среды продуктов в вещества, которые способны усваиваться организмом.
Регуляция слюноотделения. Секреция слюнных желез возбуждается рефлекторно. Пищевые или отвергаемые вещества, попадающие в полость рта и раздражающие рецепторы, вызывают безусловные слюноотделительные рефлексы. Слюноотделение через небольшой (1-3 сек) латентный период продолжается в течение всего времени, пока действует раздражитель и прекращается по окончании его действия. В продолговатом мозгу в области ядер лицевого и языкоглоточного нерва лежит центр слюноотделения. При электрическом раздражении этой области возникает обильная секреция слюны.
Парасимпатическая иннервация околоушной железы осуществляется секреторными волокнами языкоглоточного нерва, подчелюстная и подъязычные железы получают их в составе chorda thympani - ветви лицевого нерва. Симпатическая иннервация слюнных желез осуществляется волокнами из верхнего шейного симпатического узла.
Перерезка этих нервов ведет к прекращению слюноотделения. Раздражение парасимпатических волокон вызывает отделение обильного количества жидкой слюны, бедной органическими веществами. Наоборот, раздражение симпатического нерва вызывает отделение очень небольшого количества слюны, содержащей много органических веществ и ферментов.
Наряду с безусловными слюноотделительными рефлексами большую роль играют и условные - естественные и искусственные рефлексы. Болевые раздражения, отрицательные эмоции (страх) тормозят слюноотделение.
Механизмы регуляции образования и выделения желудочной слизи достаточно сложны. Оказалось, что слизь секретируется непрерывно. Стимуляция вагуса усиливает выделение слизи в желудке. Так же действуют ацетилхолин и инсулин. Особенно сильно образуется слизь в ответ на местное механическое раздражение слизистой. Процесс образования гранул мукоидного секрета стимулируется чревными нервами. В регуляции образования и выделения слизи принимают участие гистамин, гастрин, гормоны системы гипофиз - надпочечники. Адренокортикальная гиперфункция стимулирует выделение мукоида. При достаточно продолжительном воздействии наступает истощение запасов слизистых веществ в эпителиальных клетках, что приводит к нарушению защитных свойств слизистого барьера и образованию т. н. стероидных язв. Кортизон снижает концентрацию серотонина, а так как серотонин увеличивает секрецию слизи, уменьшает выход свободной кислоты и объем сока, то это тоже является одной и причин нарушения защитных свойств слизистого барьера при стрессе.
Стимуляторы и ингибиторы секреции желудочных желез. Функциональные приспособления.
Адекватными возбудителями желудочных желез являются вещества, употребляемые в пищу в сыром или переработанном виде. Желудок структурно и функционально приспособлен к такому характеру питания.
Характер желудочной секреции (непрерывная или прерывистая) зависит от условий питания. Непрерывное отделение сока наблюдается, например, у жвачных и грызунов, желудок которых никогда не опорожняется полностью. Прерывистая секреция, возникающая после приемов пищи и отсутствующая в межпищеварительном периоде, характерна для плотоядных.
На определенный порядок приема пищи вырабатывается устойчивый стереотип секреторной реакции, который при переводе на другой пищевой режим меняется, для чего требуется несколько дней.
Большинство исследователей считает, что умеренное количество сока выделяется у человека более или менее непрерывно, но в ночные и утренние часы секреция свободного от пищи желудка становится минимальной или прекращается.
Существуют разнообразные виды адаптации желудочных желез к характеру питания - видовые, индивидуальные и т. п. Классическим примером приспособительных реакций являются изученные Павловым кривые секреции сока на хлеб, мясо, молоко. Приспособительный эффект после еды различных по количеству и составу пищевых веществ достигается несколькими механизмами. Начальная адаптация происходит уже при раздражении дистантных рецепторов и рецепторов полости рта, как установили опыты с дразнением пищей и мнимым кормлением.
Основным же фактором является химический состав пищи, приходящей в соприкосновение с рецепторами привратника и кишечника. Самым эффективным возбудителем секреции желудка служит белковая пища животного и растительного происхождения, причем наблюдается тесная корреляция между содержанием белка в пище и количеством выделившейся HCl. Жир угнетает желудочную секрецию, если его 12-14% в пище. Торможение возникает при контакте жиров со слизистой 12-перстной и тонкой кишки. Углеводы слабо возбуждают кислую секрецию, а в больших количествах могут ее тормозить. Имеют значение и физико-химические свойства пищи (осмотическое давление, ее эмульгированность и т. д.).
Помимо адаптации к пище желудок участвует ряде приспособительных реакций, направленных на поддержание гомеостаза. Эти реакции вызываются не пищевыми влияниями, исходящими из внутренней и внешней среды (сезонные, климатические колебания, беременность, лактация, голодание, гипоксия, физическая работа и др.).
Эмоции оказывают неоднородное влияние на секреторную активность желудка. Астенические отрицательные эмоции (страх, печаль) ведут к его гипофункции. При состояниях возбуждения, вызываемых враждебностью, возмущением (стенические отрицательные эмоции - гнев, негодование), наоборот, наблюдается гиперфункция. Отделение сока сокращается при болевых раздражениях. Вообще любые чрезмерные по силе раздражения тормозят желудочную секрецию.
В клинических и экспериментальных исследованиях наряду с пищевыми стимуляторами (мясной бульон, капустный отвар) очень часто применяют не пищевые - алкоголь 5-10%, кофеин и др.
Регуляция выделения желудочного сока.
В зависимости от причин возникновение секреции кислого желудочного содержимого делят на 2 периода - межпищеварительный и пищеварительный. Последний в свою очередь подразделяется на 3 фазы - мозговую, желудочную и кишечную (соответ-ственно расположению чувствительных образований, с которыми взаимодействуют пищевые вещества.
Такое разделение секреции желудка на фазы возможно только в эксперименте. В естественных условиях пища попадает в желудок уже во время еды, а вскоре переходит и в кишечник. Следовательно, при акте еды раздражаются все 3, а после ее окончания - 2 воспринимающие поверхности, т. е. фазы желудочной секреции взаимно перекрываются.
Мозговая (сложно – рефлекторная, психическая) фаза развивается при раздражении обонятельных, зрительных, слуховых и др. рецепторов, когда пища находится еще вне организма, а затем механо - и хеморецепторов полости рта, глотки и пищевода. Эта фаза изучается в опытах с поддразниванием пищей или мнимым кормлением. Появление кислой реакции в желудке на 4,5-10 минут отстает от момента еды. После кратковременного раздражения секреция длится несколько часов (после 5-минутного мнимого кормления - 2-4 часа). Она легко затормаживается посторонними внешними и внутренними раздражителями, в частности эмоциональными и болевыми. Выключение из контакта с пищей перечисленных чувствительных областей в опытах с введением пищевых веществ прямо в желудок через фистулу значительно снижает и отдаляет продукцию соляной кислоты и изменяет ход секреции сока.
Желудочная (нейрогуморальная) фаза наступает при соприкосновении пищи со слизистой оболочкой самого желудка. Среди естественных контактных стимуляторов желудочных желез наиболее эффективны водные экстракты мяса и печени, а также аминокислоты и спирты. Единственной эффективной формой механического раздражения желудка является его растяжение, особенно в области привратника. Гуморальные компоненты обусловлены частичным всасыванием в желудке и отстают по времени от нервно-рефлекторного.
Кишечная (гуморально - химическая) фаза желудочной секреции возникает при переходе пищевых веществ из желудка в кишечник. Здесь происходит раздражение хемо, термо и механорецепторов кишечника и всасывание веществ. Особенностями кишечной фазы является длительный латентный период (1-3 часа), большая продолжительность секреции. Она составляет до 18% выработанного желудочного сока за весь секреторный период.
Кишечник является метом возникновения как стимулирующих, так и тормозных влияний на секрецию желудка. Секреция возбуждается продуктами белкового переваривания, молоком, алкоголем и механическими раздражениями, угнетается жирами, растворами кислот, гипертоническими (более 275 мосм) и гипотоническими (менее 50 мосм.) растворами сахаров и солей. Тормозящие влияния особенно зависят от рецепторов 12-перстной кишки. Жир оказывает тормозящее влияние на всем протяжении тонкого кишечника.
Секреторным нервом желудочных желез является вагус. Он может оказывать прямое активирующее действие на секреторные клетки (ацетилхолином), а также опосредованно - за счет стимуляции выделения гистамина и гастрина.
Адренэргические волокна тормозят кислую желудочную секрецию.
Гуморальное возбуждение и торможение деятельности желудочных желез осуществляется биогенными веществами различной структуры: пищеварительными гормонами, которые образуются в слизистой желудка и 12-перстной кишки, и так называемыми местными гормонами (гистамином, ацетилхолином), которые присутствуют в слизистой самой фундальной области. Другую группу гуморальных возбудителей желудочной секреции составляют пищевые вещества и продукты их расщепления.
Образование гормонов в пищеварительном канале находится под контролем нервной системы. Механизм их влияния на обкладочные клетки - прямой и непрямой (через гастрин).
Гастрин - гормон, стимулирующий образование HCl обкладочными клетками, вырабатывается преимущественно слизистой антрума и в меньшей степени слизистой верхних отделов желудка. Он выделяется в ответ на механические и химические воздействия на слизистую antrum piloris. Действует непосредственно на обкладочные клетки или через гистамин и АХ.
Гистамин - сосредоточен в области шеек фундальных желез. В слизистой желудка нет или мало гистаминазы. Обладает прямым стимулирующим действием на обкладочные клетки.
Гастрон - этот термин объединяет группу веществ, выделяемых из ЖКТ и способных угнетать секрецию HCl. В Эту группу кроме желудочного гастрона включаются секретин, глюкагон, энтерогастрон. Последний вырабатывается верхними отделами тонкого кишечника при воздействии жиров, гипотонических растворов и т. п. Желудочный гастрон вырабатывается привратником. Тормозной эффект зависит как от торможения образования гастрина, так и от непосредственного влияния на обкладочные клетки.
В числе гормонов 12-перстной кишки, угнетающих секрецию HCl при попадании кислого содержимого в кишку, следует назвать бульбогастрон и холецистокинин, действующие как ингибиторы гастрина.
Стероидные гормоны регулируют возбудимость желудочных желез, количество обкладочных клеток и образование слизи.
Двигательная деятельность желудка и механизмы ее регуляции.
Двигательная деятельность желудка обеспечивает прием пищи из ротовой полости, накопление ее (резервуарная функция), механическую обработку (моторная функция: перемешивание и размельчение) и продвижение содержимого в 12-перстную кишку (эвакуаторная функция). При этом желудок приспосабливается к изменению объема содержимого, не развивая чрезмерного давления и сохраняя его на определенном уровне. Указанные функции обеспечиваются деятельностью мышечной оболочки и механизмами ее регуляции.
Мышечная оболочка желудка состоит из 3-х слоев гладких мышц (продольного, циркулярного и косого). По своей структуре косой слой напоминает проводящую систему сердца. Существует мнение, что он участвует в проведении возбуждения в желудке и осуществлении координации моторной деятельности. Иннервация мышечной оболочек обеспечивается экстрамуральными нервами (вагус, чревный, диафрагмальный) и интрамуральной нервной системой.
Типы сокращений желудка. Пустой желудок человека имеет объем около 50 мл. Его полость заполнена воздухом и давление в ней соответствует внутрибрюшинному (2-6 см. Н2О). У человека прием 1 л пищи повышает давление в желудке до 7-8, максимум до 10 см. Пластический тонус желудка позволяет принимать пищу различных объемов без существенного изменения базального давления в органе.
Пустому желудку свойственна периодическая моторная деятельность. Во время расслабления базальное давление в фундальном отделе пустого желудка составляет 3-5 см Н2О, во время каждого сокращения - 30-50 см, иногда 60-100 см Н2О. Короткие периоды работы (10-30 мин), состоящие из ряда сильных сокращений, сменяются периодами покоя длительностью до 1,5 часов. Периодические сокращения желудка прекращаются с момента кормления и наступает пищевая моторика.
Выделяют три типа сокращений желудка (Богач и Гройсман - перистальтические сокращения, тонические сокращения и тонические волны Моторная деятельность желудка в процессе пищеварения зависит от вида пищи. Спустя 3-15 минут после кормления белковой или углеводной пищей в пилорическом и фундальном отделах возникают сравнительно сильные сокращения. При пище грубой консистенции (хлеб и мясо в кусках) амплитуда больше, чем при поедании измельченной пищи. Эта первая фаза пищевой моторики длится 1,5-2 часов. Вторая фаза характеризуется падением амплитуды сокращений. Появляются паузы.
Изменения силы сокращения желудка в процессе пищеварения возникают в результате взаимодействия раздражающего действия пищи, возбуждающего моторику, и тормозящего влияния из 12-перстной кишки, которое возникает после поступления туда первых порций химуса.
Механизмы регуляции моторной деятельности желудка. Моторика желудка регулируется миогенными, нервными (интра и экстрамуральными) и гуморальными механизмами. Наиболее важная роль принадлежит нервной системе.
Миогенные механизмы. Перистальтические сокращения регулируются датчиком ритма (пейсмеккером), который работает автоматически и генерирует медленные электрические волны, распространяющиеся по мускулатуре желудка.
Нервные механизмы. Основными моторными нервами желудка считаются блуждающие, тормозными - чревные. В последнее время показано, однако, что вагус в ряде случаев может оказывать тормозное влияние, эффект зависит от состояния желудка.
Гуморальные механизмы. Гормоны пищеварительного тракта и желез внутренней секреции (вазопрессин, тироксин, половые), а также физиологически активные вещества (серотонин, гистамин, брадикинин) оказывают разнообразное влияние на деятельность мышечных клеток желудка. Нервные влияния на моторику также осуществляются посредством ацетилхолина и адреналина. Результирующий эффект представляет собой алгебраическую сумму действия различных гуморальных и нервных агентов.
Рефлекторная регуляция моторики. Регуляция моторики желудка осуществляется посредством рефлексов, начинающихся в ротовой полости, глотке, пищеводе, самом желудке и различных отделах кишечника. Кроме того, рефлекторные влияния происходят при раздражении эстеро - и интерорецепторов других органов. При этом гуморальные и миогенные факторы создают фон, на котором действуют нервные механизмы.
Во время прима пищи происходит активное рефлекторное расслабление желудка. Импульсация идет с рецепторов рта, глотки, пищевода и передается по вагусам. Местные механические раздражения пищей или баллоном вызывают моторную деятельность желудка. Эта реакция осуществляется через местные интрамуральные и длинные (с замыканием в ЦНС) рефлекторные пути.
Моторная деятельность желудка подчиняется основному закону рефлекторной регуляции ЖКТ, сущность которого в том, что адекватное раздражение любого участка вызывает возбуждение моторики в данном и ниже лежащих участках, и одновременное торможение моторики выше лежащих участков ЖКТ.
Центры регуляции моторики лежат на разных уровнях ЦНС - спинной мозг, продолговатый, гипоталамус, лимбическая система, кора. Передние и средние ядра гипоталамуса преимущественно возбуждают, а задние тормозят моторную деятельность желудка. Кора оказывает условно рефлекторное воздействие.
Состав и свойства поджелудочного сока.
Выделяемый поджелудочной железой сок представляет собой прозрачную жидкость щелочной реакции (рН 7,8-8,4) , которая обусловлена наличием в соке бикарбонатов. Сок богат ферментами. В нем находятся трипсин, химотрипсин, карбоксиполипетидза, аминополипептидаза, липаза, амилаза, мальтаза, лактаза, нуклеазы и др.
Секреция поджелудочного сока начинается через 2-3 минуты после приема пищи и продолжается 6-14 часов в зависимости от состава пищи. Натощак поджелудочный сок выделяется только в небольших количествах во время периодической деятельности пищеварительного тракта. Количество сока и его ферментный состав зависят от качества поступающего химуса.
Регуляция панкреатической секреции
Регуляция выдкеления секрета поджелудочной железы осуществляется нервными и гуморальными механизмами. Секреторным нервом является вагус. Раздражение его вызывает выделение поджелудочного сока с высокой ферментной активностью. Симпатический нерв тормозит секрецию поджелудочного сока.
Рефлекторными возбудителями отделения поджелудочного сока являются раздражение вкусовых и обонятельных рецепторов, жевание, глотание. Отчетливы и условно рефлекторные воздействия.
Гуморальная регуляция секреции поджелудочной железы осуществляется благодаря образованию клетками 12-п. кишки и пилорической части желудка нескольких гормонов, которые активизируют секрецию поджелудочной железы. В 1902 г. был открыт секретин (Бейлис и Старлинг). Это был вообще первый открытый гормон. Секретин образуется из неактивного просекретина при воздействии на слизистую кишки кислоты, пептонов и гипертонических растворов. В результате поступление кислого желудочного содержимого в 12-п. кшку является мощным раздражителем секреции поджелудочной железы. Интенсивность секреторной реакции клеток поджелудочной железы а введение секретина регулируется нервной системой.
В составе экстрактов 12-п. кишки помимо секретина обнаружен и панкреозимин, который стимулирует ферментообразование поджелудочной железы. Раздражителями, вызывающими образование панкреозимина, являются пептоны, аминокислоты, жиры и жирные кислоты.
Показано что при гуморальной стимуляции выделяется преимущественно неактивный трипсиноген, а при раздражении вагуса - активный трипсин, способный переваривать белки без предварительной активации его энтерокиназой.
Рефлекторный механизм секреции поджелудочного сока - такой же, как и желудочного. Выделяют две фазы секреции поджелудочного сока - мозговую (сложно - рефлекторную) и кишечную (нейрогуморальную).
Влияние нервной системы на желчевыделительный аппарат осуществляется через блуждающие и симпатические нервы. Под влиянием импульсов, притекающих по этим нервам, происходит открытие или закрытие сфинктера общего желчного протока и сокращение или расслабление желчного пузыря. Слабое раздражение вагуса вызывает расслабление сфинктера общего желчного протока и сокращение пузыря, сильное раздражение вагуса вызывает обратный эффект.
В слизистой оболочке 12-п. кишки под влиянием продуктов переваривания белов и жиров образуется особый химический возбудитель движений желчного пузыря - холецистокинин. Он усиливает сокращения желчного пузыря и вызывает его опорожнение в разгар пищеварения. Применяется в клинике. В последнее время выяснилось, что он аналогичен ранее описанному панкреозимину.
Пищеварение в тонких кишках
Вдоль всей слизистой оболочки тонкой кишки заложены либеркюновы железы, выделяющие кишечный сок, который своим действием дополняет переваривающее действие желудочного и поджелудочного сока. Кишечный сок представляет собой бесцветную жидкость, мутную от примеси слизи, эпителиальных клеток, кристаллов холестерина. Этот сок содержит хлористый натрий и небольшое количество углекислых солей, имеет щелочную реакцию.
Кроме энтерокиназы, в кишечном соке содержатся ферменты протеолитические (карбоксиполипептидаза, аминополипептидаза, дипетидаза и др.), нуклеазы, липазы, амилаза, мальтаза, инвертаза, лактаза, кислая и щелочная фосфатазы и др. Этот комплекс ферментов ранее носил название эрепсин. Ферменты кишечного сока способны расщеплять любые вещества пищи до конечных продуктов, но особенно хорошо они действуют не на целые молекулы, а на их осколки.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 |
