

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
БИЛЕТ 23
•Спирты и кислоты, входящие в состав липидов. Кетоновые тела: биологическое значение, структура, химизм образования и окисления.
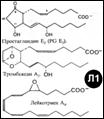 [Л1] Жирные к-ты (есть ω-номенклатура – нумерация с посл.[ω]-атома С): 1.Насыщенные (например, миристиновая – 14 атомов С, пальмитиновая 16, стеариновая 18); 2.Ненасыщенные: ·мононенасыщ. (пальмитолеиновая С16, связь 9=10; олеиновая С18, 9=10); ·полиненасыщ. (линолевая 18С, 9=10, 12=13; арахидоновая 20С, 5=6, 8=9, 11=12, 14=15); ·эйкозаноиды (производные эйкоза-20С-полиеновых ЖК): - Простаноиды (простагландины, тромбоксаны и простациклины) и –лейкотриены.
[Л1] Жирные к-ты (есть ω-номенклатура – нумерация с посл.[ω]-атома С): 1.Насыщенные (например, миристиновая – 14 атомов С, пальмитиновая 16, стеариновая 18); 2.Ненасыщенные: ·мононенасыщ. (пальмитолеиновая С16, связь 9=10; олеиновая С18, 9=10); ·полиненасыщ. (линолевая 18С, 9=10, 12=13; арахидоновая 20С, 5=6, 8=9, 11=12, 14=15); ·эйкозаноиды (производные эйкоза-20С-полиеновых ЖК): - Простаноиды (простагландины, тромбоксаны и простациклины) и –лейкотриены.
Почти все природные ненасыщ. ЖК – в цис-конфигурации.
Спирты: глицерол, холестерол, высшие спирты (цетиловый – C16H33OH, долихол)
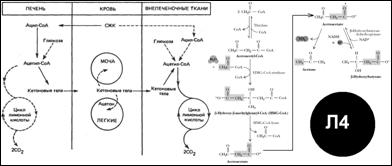
Кетоновые тела. Быстрое β-окисление ЖК: ацил-СоА ®ацетоацетил-СоА ®ацетоацетат ®ацетон или D(-)-3-гидроксибутират. Последняя р-ции катал-ся 3-гидроксибутиратдегидрогеназой (равновесие регулируется отношением [NAD+]/[NADН], т. е. окисл.-восст. статусом). [Л4]
Определяем понятия “Кетонемия”, “кетонурия”.
При голодании – исчерпание углеводных ресурсов ®усиление окисление ЖК ®накопление кетоновых тел.
Сахарный диабет ®характ. запах ацетона изо рта.
•Использование аминокислот для синтеза пигментов, тема, нейромедиаторов, гормонов и других
физиологически активных веществ. – ОТВЕТА НЕТ!!!
 •Субстраты, источники энергии, матрица, ферменты и белки ДНК-репликативного комплекса. Синтез ДНК и фазы клеточного деления.
•Субстраты, источники энергии, матрица, ферменты и белки ДНК-репликативного комплекса. Синтез ДНК и фазы клеточного деления.
Билет№23:3:![]() Для того чтобы хранящаяся вДНК инф-ция могла быть испол-на, ее необходимо переписать(транскрибировать)в последов-ть РНК. При этом ДНК служит только матрицей, т.е. она не меняется в процессе транскрипции.Транскрибируемые послед-тиДНК, т. е. участки ДНК, кот кодируют опред-ые белки, наз-ся генами.Транскрипция осуществ-ся ДНК-зависимыми РНК-полимеразами Они действуют подобно ДНК-полимеразам, за исключ того, что включают во вновь синтезируемую цепь РНК рибонуклеотиды вместо дезоксирибонуклеотидов и не нуждаются в праймерах. Эукариотические кл обычно содер по крайней мере 3 различ типа РНК-полимераз, РНК-полимераза I катализ-ет синтез РНК с коэффициентом седиментации 45S, кот служит предшествен 3-х различ рибосомных РНК. РНК-полимеразы II синтез-т гяРНК (hnRNA),кот служат предшествен-ми мРНК и мяРНК. Наконец,РНК-полимераза III транскрибирует гены, кот кодируют тРНК (tRNA), 5S - рРНК (rRNA) и некот мяРНК. Эти РНК служат предшествен-ми функцион-х РНК, кот образ-ся в процессе созревания РНК.РНК-полимераза II ингибир-ся α-аманитином(ядом бледной поганки).Стр-ра β-глобинового гена В кач примера орган-ции небол эукариот-го гена представлена схема гена, кодирующего β-цепь гемоглобина (146 аминокислот).Этот ген сост из более чем 2000 п. о. (2 тыс. п. о.).Однако из них тол около 450 п. о. из них несут инф-цию об ам. к-тной послед-ти β-глобина. Кроме 3-х кодирующих участков(экзонов),ген вкл 2 некодирующих (интроны, I1 и I2).На 5'-конце гена распол-ся промоторный участок (розовый цвет)длиной прибл-но 200 п. о., кот им регуляторную ф-цию. Транскрипция начин-ся с 3'-конца промоторн участка и продол-ся, пока не достигнет сайта полиаденилиров-я.Образ-ся первичный транскрипт (гяРНК)β-глобинового гена сост из ~1600 п. о.Во вр созрев-я РНК некодир-щие последов-сти, соответств-ие интронам, удаляются и, кроме того, оба конца гяРНК модифицир-ся. Зрелая β-глобиновая мРНК включ в себе ~40% гяРНК и дополнит-но 3'-концевую последов-ть из 100-200 АМФ (AMP).У мн генов разделение на экзоны и интроны еще более выражено. Так, общая длина гена дигидрофолатредуктазы состав выше 30 тыс. п.о. Информация об ам. к-ной послед-ти распределена по 6 экзонам, кот в сумме состав-ют только ~6 тыс. п.о.Процесс транскрипции:Как уже упоминалось,РНК-полимераза II связыв-ся с 3'-концом промоторного участка. Последов-ть, обеспеч-щая это связыв-е, так назыв-ый ТАТА-бокс,корот А - и Т-обогащенный участок, последов-ть кот слегка варьирует у разных генов.Типичная последов-ть (каноническая) — ...ТАТААА... . Для взаимод-я полимеразы с этим участком необход нескол белков,основных факторов транскрипции.Дополнит-ые факторы могут либо стимул-ть, либо ингибир-ть этот процесс (контроль транскрипции).После инициации синтеза,РНК-полимераза движется в направ-ии 3'→5' матричной цепи. В проц инициации фермент разделяет корот участок двойной спиралиДНК на 2 отдельные цепочки. Нуклеозидтрифосфаты связ-тся комплементарно на кодирующей цепочкеДНК водородными св и шаг за шагом присоед-ся к растущей молекуле РНК. Вскоре после начала элонгации 5'-конец транскрипта защищается «кэпом».Как только транскрипция доходит до сайта полиаденилирования(обычно это последов-ть ...ААТААА...),транскрипт отщепляется. После этого полимераза прекращ транскрипцию и диссоциирует от ДНК.
Для того чтобы хранящаяся вДНК инф-ция могла быть испол-на, ее необходимо переписать(транскрибировать)в последов-ть РНК. При этом ДНК служит только матрицей, т.е. она не меняется в процессе транскрипции.Транскрибируемые послед-тиДНК, т. е. участки ДНК, кот кодируют опред-ые белки, наз-ся генами.Транскрипция осуществ-ся ДНК-зависимыми РНК-полимеразами Они действуют подобно ДНК-полимеразам, за исключ того, что включают во вновь синтезируемую цепь РНК рибонуклеотиды вместо дезоксирибонуклеотидов и не нуждаются в праймерах. Эукариотические кл обычно содер по крайней мере 3 различ типа РНК-полимераз, РНК-полимераза I катализ-ет синтез РНК с коэффициентом седиментации 45S, кот служит предшествен 3-х различ рибосомных РНК. РНК-полимеразы II синтез-т гяРНК (hnRNA),кот служат предшествен-ми мРНК и мяРНК. Наконец,РНК-полимераза III транскрибирует гены, кот кодируют тРНК (tRNA), 5S - рРНК (rRNA) и некот мяРНК. Эти РНК служат предшествен-ми функцион-х РНК, кот образ-ся в процессе созревания РНК.РНК-полимераза II ингибир-ся α-аманитином(ядом бледной поганки).Стр-ра β-глобинового гена В кач примера орган-ции небол эукариот-го гена представлена схема гена, кодирующего β-цепь гемоглобина (146 аминокислот).Этот ген сост из более чем 2000 п. о. (2 тыс. п. о.).Однако из них тол около 450 п. о. из них несут инф-цию об ам. к-тной послед-ти β-глобина. Кроме 3-х кодирующих участков(экзонов),ген вкл 2 некодирующих (интроны, I1 и I2).На 5'-конце гена распол-ся промоторный участок (розовый цвет)длиной прибл-но 200 п. о., кот им регуляторную ф-цию. Транскрипция начин-ся с 3'-конца промоторн участка и продол-ся, пока не достигнет сайта полиаденилиров-я.Образ-ся первичный транскрипт (гяРНК)β-глобинового гена сост из ~1600 п. о.Во вр созрев-я РНК некодир-щие последов-сти, соответств-ие интронам, удаляются и, кроме того, оба конца гяРНК модифицир-ся. Зрелая β-глобиновая мРНК включ в себе ~40% гяРНК и дополнит-но 3'-концевую последов-ть из 100-200 АМФ (AMP).У мн генов разделение на экзоны и интроны еще более выражено. Так, общая длина гена дигидрофолатредуктазы состав выше 30 тыс. п.о. Информация об ам. к-ной послед-ти распределена по 6 экзонам, кот в сумме состав-ют только ~6 тыс. п.о.Процесс транскрипции:Как уже упоминалось,РНК-полимераза II связыв-ся с 3'-концом промоторного участка. Последов-ть, обеспеч-щая это связыв-е, так назыв-ый ТАТА-бокс,корот А - и Т-обогащенный участок, последов-ть кот слегка варьирует у разных генов.Типичная последов-ть (каноническая) — ...ТАТААА... . Для взаимод-я полимеразы с этим участком необход нескол белков,основных факторов транскрипции.Дополнит-ые факторы могут либо стимул-ть, либо ингибир-ть этот процесс (контроль транскрипции).После инициации синтеза,РНК-полимераза движется в направ-ии 3'→5' матричной цепи. В проц инициации фермент разделяет корот участок двойной спиралиДНК на 2 отдельные цепочки. Нуклеозидтрифосфаты связ-тся комплементарно на кодирующей цепочкеДНК водородными св и шаг за шагом присоед-ся к растущей молекуле РНК. Вскоре после начала элонгации 5'-конец транскрипта защищается «кэпом».Как только транскрипция доходит до сайта полиаденилирования(обычно это последов-ть ...ААТААА...),транскрипт отщепляется. После этого полимераза прекращ транскрипцию и диссоциирует от ДНК.
