

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
2.1. Опорно-двигательный аппарат
Морфологической основой движения человека является опорно-двигательный аппарат, включающий в себя костный скелет и мышечную систему. Костный скелет включает в себя более 200 единиц. Большинство из них имеют между собой подвижное сочленение в виде суставов, однако имеются и совершенно исключающие перемещения сочленения, например соединение отдельных костей черепа и крестца. Суставные соединения имеют различное строение в зависимости от того, где они расположены и какие движения необходимы в данном сочленении. Основные виды суставов представлены в таблице 1. Кроме костей каждый сустав образован связочным аппаратом. Как правило, связки образуют суставную сумку вокруг сустава, но могут находиться и внутри суставной сумки (например, в плечевом и тазобедренных суставах). Основное назначение связочного аппарата – удержание сопрягающихся костей в нужном положении без затрат энергии. Истинные суставы (диартрозы) имеют сходную «конструкцию». Сочленовные поверхности на концах костей покрыты гладким гиалиновым хрящом. И хотя хрящ – это живая ткань, в ней нет нервов и кровеносных сосудов. Нужные ему питательные вещества и кислород диффундируют через синовиальную оболочку и синовиальную жидкость. При движении хрящ уменьшает трение между костями; благодаря своей эластичности хрящ служит также амортизатором при ударе.
Связки, окружающие сустав, образуют плотную волокнистую сумку и удерживают кости в определенном положении. Связки прикреплены к обеим костям и расположены таким образом, чтобы сустав мог выдерживать приходящуюся на него нагрузку. Внутренняя полость суставной сумки выстлана синовиальной оболочкой, которая выделяет в эту полость синовиальную жидкость. Синовиальная жидкость содержит муцин и служит смазкой для суставных поверхностей − уменьшает трение между ними. Синовиальная оболочка является водонепроницаемым барьером, предотвращающим потерю синовиальной жидкости. Таким образом, сустав фактически представляет собой самоподдерживающуюся систему.
Таблица 1
Типы и функции суставов
Тип сустава | Общая характеристика | Примеры | Функции |
Неподвижный (шов), синартроидный | Тонкий слой волокнистой соединительной ткани находится между костями, жестко фиксируя их положение | Между костями черепа; между крестцом и подвздошной костью тазового пояса; между костями тазового пояса | Обеспечивает опору тела, придает прочность или защищает нежные органы, которым противопоказаны любого рода деформации |
Частично подвижный, амфиартроидный а) эллипсовидный б) вращательный | Кости отделены друг от друга хрящевыми прокладками | Суставы между позвонками, костями запястья и предплюсны Сустав между атлантом и эпистрофеем | Кости скользят относительно друг друга, но подвижность их ограниченна. Все вместе они обеспечивают разнообразные движения, придают прочность конечности Позволяет поворачивать голову |
Подвижный (синовиальный, диартроидный) а) блоковидный б) шаровидный | Сочленовные поверхности костей покрыты хрящом и разделены синовиальной полостью, заполненной синовиальной жидкостью В движении участвует относительно небольшое число мышц К костям, образующим сустав, прикреплено множество мышц | Локтевой и коленный суставы, суставы пальцев Плечевой и тазобедренный суставы | Позволяет совершать движения в одной плоскости вокруг одной оси. Может выдерживать большие нагрузки Позволяет совершать движения во всех плоскостях и некоторое вращение. Не способен выдерживать очень большие нагрузки |
Наличие суставов определяет возможность к выполнению движений, а непосредственное выполнение двигательных актов связано с работой мышечной системы, которая также представлена большим количеством отдельных мышц (более 600), выполняющих каждая в отдельности сравнительно простые перемещения частей скелета относительно друг друга. И только совместная работа нескольких мышц или их групп дает возможность выполнить сложные движения. При этом чем сложнее движение, тем большее количество мышц будет вовлечено в работу.
Способность мышц осуществлять движение связана с их возможностью изменять свою длину. При этом силовые характеристики отдельной мышцы в определенной мере зависят от ее поперечного сечения, а скоростные параметры определяются длиной мышечных волокон. Скоростные и силовые параметры сложного движения будут кроме этого определяться количеством вовлеченных в работу мышечных групп и координацией их работы. Для того чтобы поперечно-полосатое мышечное волокно могло изменять свою длину, необходимо подать на него управляющий сигнал и затратить определенное количество энергии. Чем большее количество мышечных волокон вовлечено в работу, тем большее количество энергии необходимо.
Кратко рассмотрим механизм сокращения мышечного волокна, в основе которого лежит взаимодействие миозиновых и актиновых белковых нитей под влиянием сигналов от нервной системы и с использованием энергии АТФ (аденозинтрифосфорной кислоты).
Один грамм ткани скелетной мышцы содержит примерно 100 мг «сократительных белков» − актина (молекулярная массаДа) и миозина (молекулярная масса Да). Механизм их взаимодействия во время элементарного акта мышечного сокращения объясняет теория скользящих нитей. Сократительные белки актин и миозин образуют в миофибриллах (мышечных клетках) тонкие и толстые миофиламенты. В поперечно-полосатой мышечной ткани они располагаются параллельно друг другу внутри мышечной клетки, как показано на рисунке 1, изображающем схему крошечного участка мышечного волокна человека. Миофибриллы представляют собой сократимые пучки «нитей» (филаментов) диаметром около 1 мкм. Перегородки, называемые Z-пластинками, разделяют их на несколько саркомеров длиной примерно по 2,5 мкм. С помощью светового микроскопа в миофибриллах можно видеть регулярно чередующиеся поперечные светлые и темные полосы. Такая поперечная полосатость миофибрилл обусловлена особым взаиморасположением актиновых и миозиновых филаментов. Середину каждого саркомера занимают несколько тысяч «толстых» нитей миозина диаметром примерно по 10 нм. На обоих концах саркомера находятся около 2000 «тонких» нитей актина толщиной по 5 нм, прикрепленных к Z-пластинкам наподобие щетинок в щетке. В поляризованном свете (т. е. анизотропии) она называется А-диском. По обе стороны от А-диска находятся изотропные участки, содержащие только тонкие нити и поэтому выглядящие светлыми. Пучок лежащих в определенном порядке миозиновых нитей длиной 1,6 мкм в середине саркомера выглядит в световом микроскопе темной полосой шириной 1,6 мкм; из-за свойства двойного лучепреломления. Эти так называемые I-диски тянутся до Z-пластинок. В результате такого периодического чередования светлых и темных полос в бесчисленных саркомерах миофибриллы сердечной и скелетной мускулатуры выглядят поперечно-полосатыми.
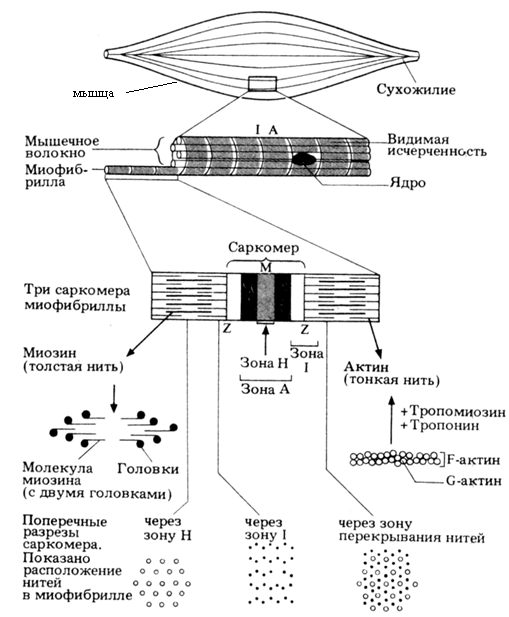
Рис. 1. Схема участка волокна скелетной мышцы человека
В покоящейся мышце концы толстых и тонких филаментов обычно лишь слабо перекрываются на границе между А и I дисками. Эта зона перекрывания в А-диске выглядит в световом микроскопе гораздо темнее центральной Н-зоны, в которой нет актиновых нитей. На электронных микрофотографиях Н-зоны видна очень тонкая темная
М-линия в середине саркомера − сеть опорных белков, по-видимому, удерживающих толстые нити в составе единого пучка.
Мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах. Сравнивая структуры саркомера в двух различных функциональных состояниях, можно видеть изменения поперечной исчерченности и взаиморасположения нитей во время сокращения: тонкие актиновые филаменты скользят вдоль толстых миозиновых, двигаясь между ними к середине их пучка.
![]()
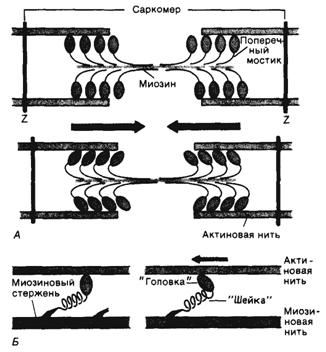 Рисунок 2 иллюстрирует основное положение теории скользящих нитей во время сокращения мышцы. Сами актиновые и миозиновые нити при сокращении мышцы не укорачиваются. Это объясняет данные световой микроскопии: ширина А-диска (1,6 мкм) всегда остается постоянной, тогда как I-диски и Н-зоны при сокращении уменьшаются.
Рисунок 2 иллюстрирует основное положение теории скользящих нитей во время сокращения мышцы. Сами актиновые и миозиновые нити при сокращении мышцы не укорачиваются. Это объясняет данные световой микроскопии: ширина А-диска (1,6 мкм) всегда остается постоянной, тогда как I-диски и Н-зоны при сокращении уменьшаются.
Длина нитей не меняется и при растяжении мышцы. Тонкие филаменты попросту вытягиваются из промежутков между толстыми нитями, так что степень перекрывания их пучков уменьшается.
Миозиновые нити несут поперечные, отходящие биполярно, как показано на рисунке 2, выступы длиной около 20 нм с головками примерно из 150 молекул миозина. Во время сокращения каждая головка (поперечный мостик) может связывать миозиновую нить с соседними актиновыми и изменять конфигурацию поперечного мостика, вследствие чего актиновые и миозиновые нити сдвигаются относительно друг друга. Движение головок создает объединенное усилие, как бы «гребок», продвигающий актиновые нити к середине саркомера. Сама биполярная организация молекул миозина обеспечивает противоположную направленность скольжения актиновых нитей в левой и правой половинах саркомера.
В результате однократного движения поперечных мостиков вдоль актиновой нити саркомер укорачивается примерно на 1% своей длины. Однако при изотоническом сокращении мышцы лягушки саркомеры за десятую долю секунды укорачиваются на 0,4 мкм, т. е. на 20% длины. Для этого поперечные мостики должны совершить свои гребковые движения за указанный промежуток времени не один, а 20 раз. Только за счет ритмичных отделений и повторных прикреплений миозиновых головок актиновая нить может подтягиваться к середине саркомера, подобно тому, как группа людей тянет длинную веревку, перебирая ее руками. Благодаря суммации минимальных укорочений миофибрилл в последовательно расположенных саркомерах мышца лягушки длиной 2 см при изотоническом сокращении за 0,1 с поднимет маленький груз на высоту 0,4 см. Следовательно, когда принцип «вытягивания веревки» реализуется во множестве последовательных саркомеров, повторяющиеся молекулярные движения поперечных мостиков приводят к макроскопическому движению. При расслаблении мышцы миозиновые головки отделяются от актиновых нитей. Поскольку актиновые и миозиновые нити могут легко скользить друг относительно друга, сопротивление расслабленных мышц растяжению очень низкое. Их можно снова растянуть до исходной длины, приложив совсем небольшое усилие. Следовательно, удлинение мышцы во время расслабления носит пассивный характер.
Благодаря упругости поперечных мостиков саркомер может развивать силу даже без скольжения нитей относительно друг друга, т. е. в строго изометрических экспериментальных условиях. Рисунок 2б иллюстрирует такой процесс генерирования изометрической силы. Сначала головка миозиновой молекулы (поперечный мостик) прикрепляется к актиновой нити под прямым углом. Затем она наклоняется под углом примерно 45° благодаря притяжению между соседними точками прикрепления на ней и на актиновой нити. При этом головка действует как миниатюрный рычаг, приводя внутреннюю упругую структуру поперечного мостика («шейки» между головкой и миозиновой нитью) в напряженное состояние. Возникающее в результате упругое растяжение достигает лишь около 10 нм. Упругое натяжение, создаваемое индивидуальным поперечным мостиком, так слабо, что для развития мышечной силы, равной 1 мН, нужно объединить усилия миллиарда таких соединенных параллельно мостиков.
Даже при изометрическом сокращении поперечные мостики не находятся непрерывно в напряженном состоянии (это наблюдается только при трупном окоченении). На самом деле каждая миозиновая головка уже через сотые или десятые доли секунды отделяется от актиновой нити; однако через такое же короткое время следует новое прикрепление к ней. Несмотря на ритмичное чередование прикреплений и отделений с частотой от 5 до 50 раз в секунду, сила, развиваемая мышечным волокном в физиологических условиях, остается неизменной, так как статистически в каждый момент времени в прикрепленном, обусловливающем напряжение состоянии находится одно и то же количество мостиков.
Мышца, поддерживающая определенное сократительное напряжение в изометрических условиях, отличается от сокращающейся изотонически тем, что с точки зрения физики не выполняет внешней работы (произведение силы на расстояние равно нулю). Однако в каждом цикле прикрепления-отделения поперечных мостиков совершается внутренняя работа по растяжению их упругих структур, которая преобразуется в тепло в момент отсоединения миозиновых головок. Теплота изометрического сокращения («изометрическая работа») за единицу времени возрастает с увеличением количества функционирующих поперечных мостиков и частоты «гребков», требующих расхода АТФ.
Чтобы выяснить роль АТФ в сокращении и расслаблении, сначала удаляли из мышечных волокон весь эндогенный АТФ (например, путем экстрагирования водным раствором глицерола, что делает мембрану проницаемой для АТФ). Такие волокна оказывались в напряженном состоянии, но при погружении в раствор АТФ вновь приобретали мягкость и растяжимость. На основании этого был сделан вывод, что АТФ расходуется для отсоединения поперечных мостиков и расслабления мышечного волокна. Для сокращения мышечного волокна необходимо присутствие в окружающей среде ионов кальция Ca++, которые выделяются из системы изолированных от цитоплазмы трубочек под влиянием электрического импульса. Сразу после выделения эти ионы снова закачиваются в полости трубочек специальными молекулами-насосами, которые и используют энергию АТФ для транспорта ионов против градиента их концентрации.
Обычно мышца возбуждается при поступлении электрических потенциалов действия от иннервирующих ее мотонейронов (нервных окончаний передающих нервные импульсы от центральной нервной системы (ЦНС) к мышечным волокнам). В результате передачи возбуждения через нервно-мышечные контакты генерируются электрические по своей природе потенциалы действия на мембране мышечной клетки (непрямая стимуляция). Под влиянием этого потенциала открываются специальные ионные каналы (представляющие собой белковые молекулы) в системе трубочек и ионы кальция выходят из трубочек в полость миофибриллы. Появление двухвалентных ионов кальция обеспечивает формирование связи между нитями актина и поперечными мостиками. В эксперименте можно вызвать мышечное сокращение, подавая непосредственно электрический сигнал на мышцу, который запустит описанную последовательность событий.
Регуляция мышечной силы в организме человека. Функциональной единицей при мышечном сокращении является двигательная единица, которая состоит из одного мотонейрона и группы иннервируемых им мышечных волокон Размеры таких единиц широко варьируют. В мышцах, выполняющих точные движения, количество мышечных волокон, иннервируемых одним мотонейроном, меньше, чем в мышцах, обеспечивающих менее точные движения. В наружных мышцах глаза, например, мотонейрон иннервирует примерно 6 волокон, в других мышцах гораздо больше, часто от 500 до 1000. Сила, развиваемая двигательной единицей при одиночном сокращении, варьирует слабо, либо все ее волокна возбуждаются и сокращаются, либо они расслаблены. Однако развиваемая сила зависит от частоты стимуляции. При высокой частоте импульсов она примерно вдвое больше, чем при низких частотах стимуляции. Даже при очень низкой ее частоте, скажем 5–10 Гц, слабое общее напряжение мышцы (тонус) остается без изменений, поскольку у разных асинхронно активных двигательных единиц максимумы одиночных сокращений не совпадают. Таким образом, несмотря на то, что мышечное волокно ритмично напрягается и расслабляется, мышца в целом может длительное время находиться в напряженном состоянии.
Нервная регуляция позы и движений. Нервная регуляция работы скелетных мышц, т. е. (регуляция запуска и выполнения всех движений), осуществляется двигательными центрами ЦНС (центральная нервная система). Они должны гарантировать строго необходимую степень возбуждения и торможения иннервирующих эти мышцы мотонейронов, чтобы возникающие мышечные сокращения обеспечивали только нужное движение. Однако точное выполнение движений возможно лишь в случае адекватного исходного положения туловища и конечностей. Нервная регуляция соответствия позы и движения − одна из важнейших функций двигательных центров коры головного мозга.
Категории двигательных актов
Рефлекторные движения. Автоматическую, стереотипную, целенаправленную реакцию организма на стимул называют рефлексом. Если у лягушки с удаленным головным мозгом ущипнуть заднюю конечность, она ее отдергивает. Если положить на спину такой лягушки смоченный кислотой кусочек фильтровальной бумаги, вскоре он будет сброшен точным движением ближайшей задней конечности. Эти эксперименты демонстрируют тот факт, что управление сравнительно простыми движениями может осуществляться на уровне спинного мозга. С разрушением спинного мозга все рефлексы исчезают, что подтверждает положение о том, что они требуют активности центральной нервной системы.
Стереотипные реакции часто возникают у животных, включая человека, в ответ на стимулы, поступающие из окружающей среды или из самого организма. Речь идет о типе поведения, который в ходе эволюции или индивидуального развития оказался наиболее эффективным ответом на данное раздражение. Всем известно множество примеров врожденных рефлексов (безусловных, по Павлову) − роговичный, кашлевой, глотательный, реакция отдергивания и т. д., а также рефлексов, приобретенных в течение жизни (условных). Однако в большинстве случаев мы их даже не осознаем. Это рефлексы, обеспечивающие прохождение и переваривание пищи в желудке и кишечнике, непрерывно приспосабливающие кровообращение и дыхание к текущим потребностям организма, и т. д. Однако попытки описать поведение человека с точки зрения цепочки рефлекторных реакций фактически исключают возможность нашего сознательного выполнения двигательных действий. Чтобы объяснить осознанное поведение человека, разработана теория на основе двигательных программ, хранящихся в памяти и запускаемых нами в нужное время сознательно.
Запрограммированные (автоматические) движения. В качестве примера программируемых действий можно привести опыты, выполненные на собаках с изолированным головным мозгом. Такие животные способны к ритмичным движениям, например, к почесыванию спины задней лапой или шаганию даже после отключения всех сенсорных входов спинного мозга путем перерезки дорсальных корешков спинного мозга, передающих в ЦНС информацию от сенсорных органов. В этом случае поступление внешних сигналов в ЦНС исключено. Следовательно, организация движений не всегда основана на рефлексах, требующих внешнего стимула. Такие последовательности движений, поддерживаемые ЦНС без внешней стимуляции, называются «запрограммированными», или автоматическими.
После того как была обнаружена способность ЦНС к такой деятельности, быстро получила признание гипотеза, согласно которой движения регулируются в основном программами, а не рефлексами, и представление о «программной организации» ЦНС стало общепринятым. Дыхание, ходьба, чесание − все это примеры врожденных программ, к которым в течение жизни индивида добавляется множество приобретенных. Среди них есть спортивные или профессиональные навыки (гимнастические движения, печатание на машинке и т. п.), становящиеся в результате соответствующей практики почти автоматическими.
Представления о рефлекторном механизме движения положили начало различным теориям поведения, делающим упор на связи стимул-реакция, тогда как на концепции программной организации базируются школы, подчеркивающие возможность независимых действий человека от внешних стимулов (их спонтанность, произвольность, непредсказуемость). Однако переоценка какого-либо одного из этих подходов малопродуктивна. Видимо, лишь сочетание их, признание того, что центральные программы подвержены влиянию сенсорных обратных связей, наилучшим образом объясняет накопленные к настоящему времени данные.
Другой важный момент состоит в том, что значительная часть нашей мышечной деятельности направлена не на осуществление движений во внешней среде, а на принятие и поддержание позы или положения тела в пространстве. Без контроля позы со стороны двигательной системы человек беспомощно рухнет на землю. Двигательная система управляет всеми целенаправленными движениями тела во внешнем мире. Они всегда сопровождаются работой и реакциями позных механизмов, идет ли речь о подготовке к движению или о коррекции позы во время или после него. Тесная взаимосвязь между позными и целенаправленными функциями − фундаментальное свойство двигательной системы. Однако полезно рассмотреть их по отдельности; смысл этого станет ясен при обсуждении роли и центральной организации различных двигательных центров, одни из которых регулируют в основном позу, а другие – целенаправленные движения.
Локализация и функции двигательных центров. Структуры, обеспечивающие нервную регуляцию позы и движения («двигательные центры»), распределены по всей центральной нервной системе от коры больших полушарий до спинного мозга. Их иерархия совершенно очевидна; это результат постепенной эволюционной адаптации двигательной системы к выполнению все более сложных задач. В процессе эволюции развитие происходит путем не столько преобразования уже существующих, сколько формирования добавочных регулирующих механизмов для выполнения новых видов деятельности. Параллельно этому повышается и специализация отдельных двигательных центров. В результате центры регуляции двигательной активности не только составляют элементы иерархической системы, но одновременно и во все возрастающей степени действуют как партнеры. Рисунок 3 схематически обобщает функции центральной нервной системы в ходе управления позой и движениями. Справа перечислены двигательные центры, слева указан их предполагаемый вклад в результирующий двигательный акт. Чтобы облегчить ориентацию в схеме, сделан упор на ее иерархическую организацию, а некоторые из высших двигательных центров (мозжечок, базальные ганглии, двигательный отдел таламуса) для простоты пропущены.
В спинном мозге сенсорные афферентные волокна (передающие нервные импульсы в направлении от периферии к ЦНС) образуют множество связей с мотонейронами, главным образом через интернейроны (промежуточные нейроны). От того, какие связи задействованы, зависит активация или торможение определенных движений. Нейронные цепи, составляющие рефлекторные дуги, на которых основаны спинальные рефлексы, представляют собой конкретные анатомические образования. Однако их деятельность в значительной мере управляется другими спинномозговыми или вышележащими центрами, специфически модулирующими прохождение сигналов по тем или иным рефлекторным дугам. Так, например, для координации работы мышц антагонистов на одной конечности достаточно нескольких нейронов в пределах одного спинномозгового сегмента. Для координации работы рук и ног человека необходимо участие разных спинномозговых сегментов. Чем сложнее по своей координации движение, тем более высоких уровней нервной системы оно требует. Такое положение сложилось в процессе эволюции. При необходимости обеспечить новые движения появлялись новые отделы центральной нервной системы, а функционирование уже имеющихся сохранялось и подчинялось управлению со стороны вновь образованных.


Рис. 3. Схема организации двигательной системы
Сам термин «рефлекс» подчеркивает, что каждое рефлекторное движение стереотипно и возникает в результате сенсорного раздражения. Однако такое определение следует расширить с учетом центральной модуляции и тормозных эффектов. Под спинальным рефлексом правильнее понимать изменение нейронной активности, вызываемое спинальными чувствительными волокнами и приводящее к запуску или торможению движения. Такие рефлексы составляют как бы «библиотеку» элементарных позных и двигательных программ, которые могут в широком диапазоне модифицироваться, интегрируясь в преднамеренное движение. Организм использует нужные программы, не привлекая высшие нервные центры к разработке деталей их выполнения. Для того чтобы мы могли ходить, нет необходимости задумываться над тем, какую мышцу и в какое время мы должны использовать. Все эти данные хранятся в той или иной двигательной программе.
Высшие двигательные системы. В эту категорию входят все центры расположенные выше спинного мозга и участвующие в двигательной регуляции. Позные функции и их координация с целенаправленными движениями контролируются главным образом структурами ствола мозга, а сами целенаправленные движения требуют участия центров еще более высоких уровней. Как показывает рисунок 5, побуждение к действию и стратегия движения формируются в подкорковых мотивационных областях и ассоциативной коре головного мозга, затем преобразуются в программы движения, те передаются в спинной мозг, а оттуда сигналы передаются к скелетным мышцам для реализации.
Сопровождающие движения типа взмахов рук при ходьбе или жестикуляции при разговоре во многих случаях контролируются подкорковыми структурами и не требуют обязательного участия двигательной коры. Исчезновение таких сопровождающих движений характерно для некоторых заболеваний, например паркинсонизма.
Взаимодействие между сенсорными и двигательными системами. Как следует из схем, представленных на рисунке 3, сенсорная информация и двигательная активность тесно взаимосвязаны. Для правильного выполнения движений необходимо, чтобы ко всем отвечающим за это структурам в каждый момент времени поступала с периферии информация о положении тела и о ходе реализации составленной программы. Кроме того, некоторые виды сенсорной информации (например, зрительную и тактильную) можно получить только с помощью определенных двигательных актов. В обоих случаях некоторая часть сенсорных сигналов достигает двигательной системы коротким локальным путем (например, посредством спинальных рефлексов), а остальные идут по длинным петлям, захватывающим, по всей вероятности, корковые структуры.
Для обеспечения ЦНС информацией о состоянии мышц и связок служат специализированные клетки. В мышцах расположены мышечные веретена, представляющие собой соединительнотканную капсулу, которая окружает группу мышечных волокон, тоньше и короче обычных. Те, что заключены в капсулу, называются интрафузальными. Все прочие, составляющие основную массу мышцы и обеспечивающие ее работу − экстрафузальными. Различия в их размерах весьма значительны: диаметр первых 15–30 мкм, длина 4−7 мм, тогда как у вторых диаметр примерно 50−100 мкм и длина от нескольких миллиметров до многих сантиметров. Каждым своим концом мышечное веретено прикрепляется к соединительнотканной оболочке пучка экстрафузальных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5–1 мм. Мышечные веретена реагируют на растяжение, посылая нервные импульсы в ЦНС.
У наземных позвоночных в сухожилиях всех мышц, вблизи от сухожильно-мышечного соединения, находятся особые рецепторы − сухожильные органы (или сухожильные органы Гольджи). Они состоят из сухожильных нитей, отходящих примерно от 10 экстрафузальных мышечных волокон и заключенных в соединительнотканную капсулу. Они, так же, как и мышечные веретена, реагируют на растяжение.
Мышечные веретена есть практически во всех поперечнополосатых мышцах млекопитающих. Количество мышечных веретен в каждой мышце зависит от ее размера и функции. У человека их от 40 (в мелких мышцах кисти) до 500 (в трехглавой мышце плеча), а во всей мускулатуре околоПлотность мышечных веретен, т. е. их число на 1 г мышечной ткани, особенно высока в мелких мышцах, участвующих в выполнении тонких движений, например в мелких мышцах кисти (до 130); в крупных мышцах туловища это число значительно меньше. Число сухожильных органов для разных мышц до сих пор подробно не изучено. По очень приблизительным подсчетам, их 50–80 на каждые 100 мышечных веретен.
Рецепторные функции мышечных веретен и сухожильных органов. По типу адекватного раздражителя мышечные веретена и сухожильные органы относятся к рецепторам растяжения. Однако их расположение различно; первые лежат параллельно экстрафузальным волокнам, а вторые соединены с ними последовательно. В результате неодинаков и характер их активации, особенно во время сокращения мышцы. Когда растяжение мышцы примерно соответствует ее длине покоя, в большинстве первичных окончаний веретен регистрируются импульсы, тогда как сухожильные органы, как правило, «молчат». Во время растяжения частота импульсации в волокнах мышечных веретен возрастает и появляется активность в сухожильных органах. При изотоническом сокращении экстрафузальных мышечных волокон напряжение мышечных веретен снижается, и импульсация от них прекращается. Однако сухожильные органы при этом остаются растянутыми и частота их разряда даже кратковременно увеличивается.
Следовательно, мышечные веретена регистрируют главным образом длину мышцы, а сухожильные органы – ее напряжение. Реально процесс регуляции длины мышцы и ее напряжения включает еще несколько групп нервных волокон, которые мы для простоты не рассматриваем.
2.2. Энергообеспечение работы мышц
Для того, что бы мышца могла длительно выполнять работу, необходимо обеспечить непрерывный ресинтез АТФ с использованием энергии субстратов биологического окисления. Биологическое окисление субстратов представляет собой цепь последовательных реакций, конечными продуктами которых являются углекислый газ и вода. Поставщиками субстратов биологического окисления в организме являются белки, жиры и углеводы, которые в результате целого ряда реакций превращаются в более простые вещества, способные при окислении выделять энергию, необходимую для ресинтеза АТФ.
Субстраты для ресинтеза АТФ различны в зависимости от снабжения организма кислородом. При анаэробном (без участия кислорода) гликолитическом синтезе АТФ расщепляется мышечный гликоген, содержание которого в процессе работы уменьшается. Свободная глюкоза, приносимая кровью, используется мало, поскольку для вовлечения ее в гликолиз требуется присоединение к ней остатка фосфорной кислоты, происходящее с участием АТФ, которой недостаточно, так как она расходуется на энергетическое обеспечение мышечных сокращений. Гликоген расходуется без затруднений, поскольку для образования из него гексозофосфорных эфиров требуется лишь неорганическая фосфорная кислота, содержание которой в мышце при работе повышено вследствие расщепления АТФ. Возможности такого энергообеспечения сильно ограничены как наличием гликогена в мышцах (у человека в среднем около 350 г во всех мышцах), так и накоплением кислых продуктов реакции. Также следует отметить, что КПД такого энергообеспечения очень низкий (в 17 раз ниже, чем при окислении с участием кислорода).
При переходе к аэробному окислению расход мышечного гликогена уменьшается и все больше используется приносимая кровью глюкоза. В связи с тем, что она образуется из гликогена печени, он быстро расходуется, и его содержание уменьшается. В зависимости от интенсивности работы углеводы способны поддерживать движения от одного до полутора часов. К моменту окончания этого срока начинают окисляться источники энергии неуглеводной природы. Происходит мобилизация липидов из жировой ткани.
Основными поставщиками энергии в организме являются углеводы, но при тяжелой длительной работе в процесс вовлекаются жиры, а при их истощении и белки.
При расщеплении 1 г углеводов выделяется 4,1 ккал. Особенно интенсивно углеводы используются мышцами и клетками головного мозга. В организме постоянно имеются некоторые запасы углеводов в виде глюкозы. Так, например, содержание глюкозы в крови в норме должно находиться в пределах от 0,08 до 0,12 %. Организм обладает свойством откладывать углеводы в запас в виде гликогена в печени и в мышцах, а также превращать излишние количества углеводов в жиры. Величина запаса гликогена в среднем составляет около 350 г, а у спортсменов может достигать 500 г, что увеличивает их потенциальные возможности к проявлению как физической, так и умственной работоспособности. Перед интенсивной умственной или физической работой содержание глюкозы в крови рефлекторно повышается. При этом интересно отметить, что это повышение перед умственной работой такое же, как и перед физической. Снижение концентрации глюкозы в крови ниже уровня 0,07% снижает мышечную и умственную работоспособность, приводя к прекращению выполнения работы. Признаками подобного состояния являются учащенное дыхание, активное потоотделение, слабость, головокружение. Однако под влиянием тренировки у спортсменов иногда отмечается снижение уровня глюкозы в крови вплоть до 0,04%.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
