
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
То есть условный рефлекс должен распознать событие до появления безусловного раздражителя и среагировать раньше, как будто бы это событие уже произошло. Просто ассоциация активируется – припоминается. То есть при виде огня субъект как бы «убирает» руку из огня и отступает назад. Но затем, не получая подкрепление ожогом, рефлекс убирание руки при виде огня угасает, а условный рефлекс отступания назад подкрепляется видом огня и сохраняется. Поэтому в данной ситуации вид огня уже выступает в роли безусловного раздражителя и подкрепляет рефлекс отступания от огня.
Каков механизм такой длительной задержки? Одно из возможных объяснений сводится к тому, что врожденное чувство страха поддерживает у крысы образ незнакомой пищи в активном состоянии длительное время, и если наступают симптомы отравления, то они накладываются на активный образ и подкрепляют его.
Если исходить из того, что условный рефлекс как бы прогнозирует наступление события «А», тогда возникает вопрос, насколько далеко возможно прогнозирование с помощью условных рефлексов? Иными словами, как долго может сохраняться в памяти условный стимул, ожидая подкрепления? Тогда должен появляться сложный или составной рефлекс и на промежуток времени, или, проще говоря, на величину задержки. Каков нейрохимический механизм этой задержки? Где находится таймер? Как запоминаются интервалы времени?
Один из возможных механизмов – это наличие нейронов, спонтанно разряжающихся и объединенных в некую цепь. Например, должны существовать милисекундные нейроны, секундные, минутные, часовые и суточные (до семи суток, затем отсчет промежутков времени невозможен?). Тогда механизм задержки представляет собой установление условного рефлекса между условным стимулом и тем временным нейроном, который активен в момент прихода сигнала подкрепления.
ИМХО – не туда пошли.
Поэтому в систему введем еще одну модальность – временную. Ее сенсорные нейроны представлены множеством нейронов с разным циклом разрядной активности, причем на более высоком уровне, временной нейрон с более медленным циклом разряда подавляет нейроны с более коротким.
Разница между подкреплением поведения и классическим условным рефлексом
Подкрепление – это решение проблемы, безусловный стимул – это рождение проблемы.
Расчет количества нейронов в искусственной нейросинаптической сети
Исходя из принципа дихотомичности условного рефлекса и опираясь на предыдущие выкладки можно вывести формулы и правила для расчета архитектуры и количества элементов нейросинаптической сети. Это в конечном итоге должно нас привести к созданию универсальной топологии нейросинаптической сети, но с разным количеством нейросинаптических элементов. Количество нейросинаптических элементов рассчитывается по простым формулам исходя из универсальной топологии.
Хмы Хмы :)
Введем понятие универсальной топологии. Универсальная топология – это обобщенная топология нейронной сети, характерная для различных видов животных (от беспозвоночных и до позвоночных). Она представлена сенсорными системами, генетически детерминированными связями со вставочными нейронами, недетерминированными (химическими) связями между вставочными нейронами и ассоциативными клетками, детерминированными связями между ассоциативным слоем и мотонейронами.
Хмы, хмы :) Ох уж эта стратсь к универсалиям. Все нет их больше. Не в мозге, ни в молекулах, ни в нано. Кончились унивресальные законы в науке. Нет теперь такой халявы как яблоко. В том и сложность современного этапа науки. Прошу прощения.
Описание одного слоя элементарной синаптической сети
Пусть мы имеем две модальности с диапазоном значений от 1 до 3-х в каждой, с шагом 1, тогда сенсорных нейронов будет по 3 на каждую модальность, ассоциативный нейрон один и один выходной нейрон. Как уже упоминалось количество мотонейронов зависит от количества исполнительных устройств или «мышц».(на схеме их 5)
![]() E2
E2
 |
V3
![]()
V1 K1
S2 M1
 V2
V2
S1 E1
V1 – сенсорные нейроны двух модальностей, V2 – вставочные нейроны, S1 – химические синапсы, K1 – ассоциативный нейрон, V3 – промежуточный нейрон (выходной), S2 – химические синапсы, M1- Мотонейроны, E1, E2 – электрические синапсы по которому передается подкрепление или внесинаптическое химическое подкрепление.
Как видно из схемы, химические синапсы имеет только ассоциативный нейрон и мотонейроны, а все остальные нейроны имеют электротонические синапсы. Поэтому изменение синаптических весов происходит только в химических синапсах. При появлении на каком-либо химическом синапсе (или синапсах) сигнала и одновременном появлении сигнала в электрическом синапсе E1 или E2, происходит модификация соответствующего химического синапса. Обучение ассоциативного нейрона и мотонейронов независимое.
Конечно, тут в Вашу пользу есть данные: универсальность колонок в коре и наличие эфапсов. Вы мне потом эту схему подробнее объясните. Но думаю это все схожесть, а не универсальность.
А существует ли нейробиологический аналог вышеприведенной архитектуры? Наверное, его нужно искать в простейших нейробиологических сетях, раз мы предполагаем, что принципы передачи информации у беспозвоночных и позвоночных животных одинаковы. Описание такой сети я нашел у Кендела:
Архитектура этой сети полностью укладывается в теоретические представления об архитектуре, изложенные выше: сенсорные нейроны->вставочные нейроны (на рисунке они не показаны)->интернейроны с химическими синапсами->выходной нейрон (ЛГ) с электричесими синапсами->мотонейроны с химическими синапсами
Для того чтобы еще больше упростить графическое представление синаптической сети и сделать ее более наглядной, объединим сенсорные нейроны в два блока, а вставочные нейроны, химические синапсы и ассоциативный нейрон представим в виде нейроматрицы.
E2
 |
![]() M1
M1
![]()
![]() V3
V3
M2
 |
 V1 E1
V1 E1
На самом деле потенциальные синаптические связи между рецепторными нейронами и ассоциативными в начальный момент обучения в системе отсутствуют. Синаптическая связь образуется только при одновременном появлении сигнала на любой паре рецепторных нейронах в разных модальностях и сигнала подкрепления на ассоциативном нейроне. Это так.
Ваша модель красива и вполне будет работать.
Если взять количество сенсорных нейронов одной модальности за m1, а второй m2, тогда потенциальное количество синапсов Sn на одном ассоциативном нейроне будет Sn=m1*m2+F, где F это количество факторов подкрепления для данного ассоциативного нейрона. В простых сетях он равен 1. Фактор F может быть представлен, как подкрепляющим синапсом, так и внесинаптическим подкрепляющим фактором, действующим на все активные синапсы данного нейрона одновременно. Предсказать, где и сколько будет реально образовано синапсов на данном нейроне невозможно. Это будет зависеть от обучающей выборки.
24.02.2007г.
Колонки и микро-колонки
Микро-колонка представляет собой локальную нейросинаптическую сеть, основанную на универсальной топологии и моделирующую образование рефлекторной связи между любой парой модальностей. Согласен.
Колонка представляет собой локальную нейросинаптическую сеть, основанную на универсальной топологии и моделирующую образование рефлекторной связи между всеми модальностями. (рис.3) хороший принцип.
Колонки состоят из микро-колонок. (рис.2)
Каждая колонка имеет только один выходной нейрон.
Все колонки нейросинаптической сети имеют общие сенсорные нейроны.
Каждый выходной нейрон колонки посылает тормозящие синапсы на все выходные нейроны других колонок.
Элементарный (простой) условный рефлекс представлен в данной сети одним синапсом на ассоциативном нейроне. (рис.1)
Правило образование простого условного рефлекса: при одновременном появлении условно-рефлекторного сигнала на входном нейроне и сигнала подкрепления на ассоциативном нейроне, между ними образуется синапс. Усиливается. (прим. автора:согласен)
Вес синапса определяется суммой текущего и предыдущего веса данного синапса. ЭТО так. Это очень важно. Но надо говорить точнее: к старым значениям прибавляются новые. Например вновь синтезированные рецепторы добавляются к старым. (прим. автора: важное замечание)
Текущий вес синапса определяется частотой входного сигнала. Чем выше частота, тем выше текущий вес синапса.
Совсем не обязательно, так как дендрит значительно преобразует сигнал, уменьшает или увеличивает. (прим. автора : этот тезис был заявлен на будущее, в данной модели он не используется)
Частота входного сигнала является мерой воздействия на сенсорные нейроны. Чем сильнее воздействие, тем выше частота.
Если Вы имели в виду только активацию первого слоя, то как определяется частота входного сигнала (ком. автора: правильнее было бы сказать «частота выходного сигнала»)
Правило модификации синапса: изменение веса синапса происходит только при совпадении по времени сигнала на пресинаптическом нейроне и сигнала подкрепления на ассоциативном-постсинаптическом нейроне.
Достаточно близковременности, а не только совпадения (прим. автора: согласен, но это не меняет сути)
Правило прохождения сигнала через синапс: при отсутствии сигнала подкрепления на ассоциативном нейроне, вес сигнала, проходящего через синапс равен весу синапса и не зависит от частоты входящего сигнала.
Синаптическая суперпозиция: текущий вес выходного сигнала данного нейрона является суммой весов активных синапсов в данный момент, причем вес тормозящего синапса является отрицательной величиной.
Не обязательно, возможны функции коррекции (прим. автора: возможны…)
Правило внесинаптического подкрепления: Внесинаптическое подкрепление действует только в пределах одной колонки и разрешает модификацию весов всех активных синапсов в данный момент.
Это дискуссионное допущение, модель среды по сути, надо обсуждать (прим. автора: естественно…,возможно надо было уточнить, что в пределах данной активной колонки?)
Активным синапсом считается синапс, у которого есть сигнал на пресинаптической мембране.
Не обязательно, он может быть подавлен другими аксонамиJ) (прим. автора: полностью согласен! Здесь я погорячился…наверное надо было сказать на «постсинаптической мембране»…Хотя и первая трактовка в данной модели работаетJ)
Термины вес сигнала и частота сигнала в данной модели являются синонимами.
Нет нет их нельзя путатьJ (прим. автора: без комменариев…)
 |
 u s
u s
k
p
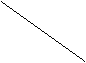
рис.1 Элементарный условный рефлекс.
u – условный стимул s – синапс, k – ассоциативный нейрон, p – сигнал подкрепления
 | |


![]()
![]()

![]()

![]()
![]()
![]() s1 k
s1 k
![]()
![]() si
si
![]() p
p
рис. 2. Микро-колонка.
![]() - вставочные нейроны, si – синапсы, k- ассоциативный нейрон, p - сигнал подкрепления
- вставочные нейроны, si – синапсы, k- ассоциативный нейрон, p - сигнал подкрепления
(прим. автора: сигнал подкрепления может быть как электрическим, так и гуморальным)
 | |
![]()
![]()

![]()
![]()
![]()

 w1 s1
w1 s1


![]()
![]()
 m1 k1
m1 k1


![]() s
s i
i
Vs V
 |

![]()
 s1
s1
![]()
![]() ki
ki
![]() p
p
mi wi si
рис.3 Колонка.
mi – сенсорные нейроны различных модальностей, Vs – суммирующий нейрон с электрическими синапсами, V - выходной нейрон.
Подключение сенсорных нейронов различных модальностей к колонке
Как видим, топологию связей в синаптической сети определяет условный рефлекс. Он же определяет и топологию связей между сенсорными нейронами различных модальностей и колонкой. Следует заметить, что в качестве «сенсорных нейронов» могут выступать и выходные нейроны предыдущих колонок.
Сначала рассмотрим типы условных рефлексов, реализованных в синаптической сети.
Вариант первый. Одна реакция при множестве условных раздражителей. Реализация возможна в одной микро-колонке.
U1+B=A
U2+B=A
Ui +B=A
 |
![]()

![]()
![]()
![]()
![]()
![]()
![]() s1 k
s1 k
![]()
![]() si
si
![]() p
p
рис.4
Вариант второй. Один условный раздражитель и множество реакций. Реализация возможна в нескольких колонках, так как множество реакций предполагает множество выходных нейронов, а в колонке он один. Вариант реализован с помощью элементарных условных рефлексов в разных колонках.
U+B1=A1
 U+B2=A2
U+B2=A2
U+Bi =Ai
 |


![]()

 |
p1
 |

![]()
![]()
 | |
 p2
p2
|
![]()
![]()
pi
рис.5
Вариант третий. Множество раздражителей – множество реакций. Реализация возможна в нескольких колонках, так как множество реакций предполагает множество выходных нейронов, а в колонке он один. Вариант реализован с помощью элементарных условных рефлексов в разных колонках.
U1+B1=A1
 U2+B2=A2
U2+B2=A2
Ui +Bi = Ai
|
![]()
![]()
 |
p1
|
![]()
![]()
| |
 p2
p2
|
![]()
![]()
![]()
pi
рис.6
Вариант четвертый. Вариант реализован с помощью микро-колонок в разных колонках.
U1+B1=A1 U1+B2=A2 U1+B3=A3

![]()
![]() U2+B1=A1 U2+B2=A2 U2+B3=A3
U2+B1=A1 U2+B2=A2 U2+B3=A3
Ui+ Bj =Aj, Ui +Bj =Aj, Ui +Bj = Aj
|
U1 ![]()
![]()
![]()
![]()

![]()
![]()
![]() s1 k1
s1 k1
![]()
![]() Ui si
Ui si
![]() p1
p1

|
U1 ![]()
![]()
![]()
![]()
![]()
![]()
![]() s1 kj
s1 kj
![]()
![]() Ui si
Ui si
![]() pj
pj
рис.7
Это красиво, но мне нужно это прояснитьJ
Теперь рассмотрим проблему подключения сенсорных нейронов к колонкам. Вы, наверное, уже заметили, что теория синаптических сетей старательно избегает полносвязных структур. Полносвязные структуры обычно возникают от бессилия понять логику и функцию связей. Это как правило так и есть В теории синаптических сетей, если Вы заметили, функция всегда определяет структуру. Йес! Особенно нейробиологические функции, которые мы пытаемся моделировать…
Мы уже рассмотрели простые условные рефлексы, теперь перейдем к более сложным рефлексам – составным. Допустим, нам необходимо отследить связь (или, что, то же самое – выработать условный рефлекс) между тремя условными стимулами разной модальности. Но мы не уверены, существует ли такая связь. Возможно, что связь существует между двумя модальностями из трех, а может быть и между всеми тремя модальностями. Из аксиомы дихотомичности условного рефлекса следует, что условный стимул всегда один. Поэтому мы должны объединить эти модальности в один условный стимул и подать его на колонку. Из этого следует, что вставочный нейрон колонки должен выполнять функцию «И». Осталось выяснить, сколько входов должно быть у этого нейрона и как объединять модальности? Обозначим модальности через M1,M2,M3. Если мы подадим на вход колонки сигнал М1^M2^M3, то мы сможем установить связь между этими сигналами и безусловным стимулом, только в том случае, если одновременно на вход колонки придут три сигнала от разных модальностей. Если же придут два сигнала, то вставочный нейрон их не пропустит и связь не будет образована. (те же рассуждения справедливы и для большего числа модальностей), поэтому разобьем модальности по парам. Из теории комбинаторики следует, что этих пар будет три – M1^M2, M1^M3, M2^M3. Исходя из теоремы 1, имеем вариант первый: одна реакция при множестве условных раздражителей. Реализуется первый вариант в одной микро-колонке:
Красиво.
M1^M2 ![]()
![]()
![]()
![]()
![]()

![]()
![]()
![]() s1 k
s1 k
![]()
![]() M1^M3
M1^M3
![]()
![]() s2
s2

![]() p
p
![]() M2^M3 s3
M2^M3 s3
![]()
![]()
рис.8
Из условия видно, что вставочный нейрон Wi должен быть двухвходовой, то есть реализовывать функцию «И» по модулю 2.
Но из филогенеза нервной системы мы знаем, что увеличение объема и усложнение структуры отделов головного мозга тесно связано с развитием сенсорных систем. Развитие отделов мозга связано с развитием той или иной сенсорной системы. К тому же и сама централизация и цефализация являются результатом развития сенсорных (чувствительных) структур. Из этого видно, что чем более развита сенсорная система, тем более сложное строение будет иметь нервная система. Отсюда недалеко до вывода о том, что чем сложнее сенсорная система в модели, тем сложнее и нейросинаптическая сеть. Но усложнение сенсорных входов может привести к проклятию размерности, которое мы намерены избежать. Если все наши предыдущие предположения были верны, то топология и структура связей нейросинаптической сети будет неизменна, а усложняться будет только архитектура этих связей при усложнении сенсорных входов. Это все равно будет требовать программно-аппартных ресурсов. (прим. автора: не торопитесь, ведь я еще ничего не сказал об алгоритме обучения и распознавания…)
Колонки, по своей локализации, находятся в корковом слое головного мозга. Между сенсорными нейронами и корковым представительством какой либо модальности сигнал претерпевает значительные изменения. Он структурируется. Выделяется его интенсивность и локализация. Между сенсорными клетками и корковой проекцией модальности имеются клетки, по-разному реагирующие на интенсивность и локализацию сигнала. Одни клетки реагируют на слабый сигнал и игнорируют сильный, другие наоборот и всегда есть клетки, реагирующие только на определенное значение и локализацию сигнала. Эти клетки обычно организованы в ядра. В них представлены клетки, реагирующие на весь диапазон значений данной модальности. То есть, независимо от типа рецепторов, в этих ядрах происходит стандартизация и структуризация входного сигнала и приведение его к виду, доступному для обработки различными корковыми структурами. Поэтому, говоря о входных «сенсорных» нейронах в данной модели, подразумеваются структуры, которые работают уже с обработанным, стандартизованным и структурированным сигналом. В анатомическом смысле это могут быть выходные нейроны ядерных структур.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
