
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Уважаемый Олаф!
С огромным интересом прочитал Ваш материал. Мне кажется у Вас очень плодотворный подход. Я еще не вполне разобрался в Вашей архитектуре колонок, но, думаю, мы еще сможем ее по обсуждать. То, что у меня вызвало вопросы или несогласие – выделил красным. Но, видимо, большая часть – это именно вопросы, быть может в силу не полного понимания особенностей Вашей архитектуры.
С уважением и наилучшими пожеланиями,
![]() 19.10.2006г. Нейробиологическое обоснование модели.
19.10.2006г. Нейробиологическое обоснование модели.
Олаф Спрэнг
Цель данной работы
Создать модель искусственной нейросинаптической сети, максимально используя нейробиологические принципы и решающую следующие задачи:
1. Универсальность сети – сеть должна быть универсальной в том смысле, что единая архитектура и алгоритм обучения должны обеспечивать решение самых разнообразных задач, связанных с классификацией, распознаванием образов, обобщением, установлением связей между различными событиями, образами, классификациями.
2. Предсказуемость результатов обучения – исходя из топологии сети, всегда можно заранее предсказать, каков будет результат ее работы исходя из условий задачи.
3. Легкость проектирования сети под конкретную задачу – сеть автоматически конфигурирует свою топологию исходя из анализа входных и выходных данных.
4. Исходя из парадигмы построения нейросинаптических сетей, всегда генерируется единственная и оптимальная для данной задачи сеть.
5. Устойчивость к искаженным входным данным – при равномерном случайном распределении ошибки во входных данных, величина ошибочных данных не может превышать 49% от всего объема обучающей выборки.
Может мы и по разному что-то представляем но эти тезисы находят подтверждение и в моих представлениях
6. Масштабируемость – предполагает отсутствие проклятия размерности. Количество синапсов и нейронов в сети ограничено только условием задачи.
Программно-аппаратные ограничения, к сожалению, все же есть (прим. автора: с этим не согласен – ограничения только в программистеJ)
Основные нейробиологические посылки
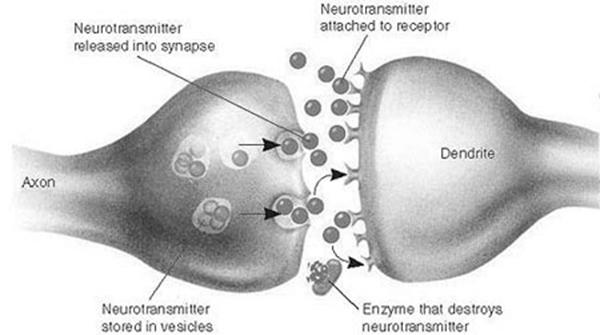
В 1943 году, когда зарождалась теория искусственных нейронных систем, синапсы еще не были хорошо изучены, поэтому за основу был взят нейрон. Но в настоящее время очевидно, что нейрон обрабатывает только часть информации, являясь по сути только генератором импульсов, управляет же нейроном синапс, разрешая или нет нейрону генерировать импульс, а также задавая частоту и силу этого импульса. Синапсы расположенные более периферийно (на дендритах) могут осуществлять более тонкий контроль возбудимости постсинаптического нейрона, чем синапсы, оканчивающиеся на теле нейрона.(С. Немечек, 1978г. с.203)
Биологический химический синапс состоит из пресинаптической мембраны, находящейся на аксоне одного нейрона и постсинаптической мембраны, находящейся на дендрите другого нейрона. В синаптическую щель между мембранами из постсинаптической мембраны при появлении сигнала на аксоне выделяется нейромедиатор, который связываясь с рецепторами на постсинаптической мембране, вызывает генерацию сигнала телом нейрона. Затем происходит быстрое разрушение нейромедиатора.
Провокационный вопрос: а есть ли синапс вообще? :)))
Ведь можно и так поставить вопрос: есть пре и постсинаптические мембраны. Они обе – части соотв. Нейронов. Собственно щель обеспечивает лишь диффузию медиатора. Изменения происходят в мембранах нейронов
Фехнер (Fechner, 1860) выдвинул предположение, что ощущение пропорционально логарифму силы раздражения. Эдриан (Adrian, 1930) обнаружил, что нейроны сигнализируют об изменении интенсивности стимула, изменяя частоту своего разряда.
Под влиянием стимула происходит целый каскад изменений нейрона: есть динамика постсинаптических потенциалов, есть синтез рецепторов, разных причем, частота ПД и результирующая фасилитация, приводящая к накоплению кальция и активации синапсина, регулирующего количество открывающихся везикул, лишь один из нескольких важных феноменов
Хартлайн и Грэм (Hartline, Graham, 1932) обнаружили, что частота разряда нейрона пропорциональна логарифму интенсивности стимула. Кэндел (Kandel, 1976) показал, что частота импульса нейрона зависит от количества квантов медиатора, выделяемого в синаптическую щель пресинаптичесой мембраной.
Нет, нет это Вы много чего пропускаете. Частота зависит от превышения порога суммой ВПСП. А та зависит от числа событий медиаторно-рецепторного соединения в постсинаптической мембране. Грубо так.
Чувствительность постсинаптической мембраны остается неизменной.
Ни в коем случае. Ведь синтезируются новые рецепторы в постсинапсе и увеличивается число везикул в пресинапсе. (прим. автора: полностю согласен. В данном случае этот тезис дан для упрощения модели)
Транскрибирующие механизмы ядра под влиянием потенциала действия обеспечивают биосинтез молекул медиатора и др. необходимых белков, а синтезированные с помощью РНК молекулы рецепторов достаточно свободно распространяются по всей клетке, как в области активированных, так и в области не активированных постсинапсов; при этом данные молекулы встроятся в постсинаптические мембраны только тех синапсов, которые претерпели временное (среднесрочное) повышение своей эффективности – т. е. только в те дендриты, которые провели данный стимул (по-видимому, факторами встраивания синтезированных молекул в мембраны играют молекулы фосфорилированные или активированные иным образом в результате входа кальция в эксордиум дендрита при активации постсинапса); (прим. автора: с этим полностью согласен)
Посмотрите
http://www. *****/2005/5/neyrobiology. shtml Розенталь (Rosenthal, 1969) нашла, что при усилении синаптической передачи запас медиатора не увеличивается, а увеличивается только вероятность его выхода в синаптическую щель.
Нет, нет это не так, количество везикул с медиатором растет, растет площадь шипиков (прим. автора: да это так…, но это просто обзор, историю науки нам с Вами не переписатьJ)
Так же было показано, что количество квантов медиатора выделяемого в синаптическую щель одного синапса зависит от активации проводящего пути другого синапса.
Я догадываюсь о какой схеме сети Вы говорите, но такая редакция высказывания может вести к принципиальной ошибке. Вы скорее всего тут говорите о треугольнике Хэбба, но там более сложный механизмы.
То есть возникает гетеросинаптическим путем. Кэндел смоделировал появление условного рефлекса, использовав два разных пути, конвергирующих на одном нейроне – это Хэбба синапс. При одновременной активации сильным безусловным стимулом одного пути и слабым условным стимулом другого пути он через некоторое время добился усиления разряда нейрона слабым условным стимулом на 100-800%. Да в результате синтеза рецепторов в первоначально слабом синапсе.
Возникающие в течении онтогенетического развития размещение и густота синапсов на рецепторной поверхности постсинаптического нейрона не является случайным. Часто наблюдается характерное расположение не только в корковых слоях, но и в остальном сером веществе.(С. Немечек, 1978г. с.201)
Естественно на то и гены.
Базовые предположения
Наиболее подходящим местом для следов памяти являются связи, или синапсы, через которые осуществляется взаимодействие отдельных пар нейронов.Не согласен. Во первых не синапсы, а постсинаптическая мембрана и пресинаптический бутон. (прим. автора: это вопрос терминологии – Вы считаете, что есть пресинаптический бутон, синаптическая щель и постсинаптическая мембрана, а я говорю синапс, подразумевая все вышесказанное. Мы же не будем, когда собираемся говорить об архитектуре связей и функциях этих связей уточнять химический состав нейромедиаторов и нейромодуляторов? Вот об этом я и говорю, что всегда необходимо четко представлять тот уровень, на котором находишься, и четко представлять иерархию этого уровня.) И это только для долговременной памяти, а это последняя стадия (прим. автора: возможно да, возможно – нет…)
Системы, в которых подкрепление относится к связям, гораздо экономичнее систем, в которых подкрепление прикладывается к целому нейрону.Основные понятия
Синапсы
Описание формального химического синапса.
Формальный синапс представляет собой регистр памяти, в которую записывается некоторое число, характеризующее количество квантов, выбрасываемых в синаптическую щель при активации. При выработке условного рефлекса число квантов медиатора растет. Число квантов медиатора, выбрасываемого в синаптическую щель при активации синапса будем считать весом данного синапса.
Растет и число медиатора и число принимающих рецепторов. Или уменьшается при забывании (прим. автора: возможно с этим я и соглашусь, но на дифференциацию веса это никак не влияет. Вес синапса, как раз, и характеризуется всеми этими факторами)
Описание формального электрического синапса.
Формальный электрический синапс представляет собой связь между нейронами. Вес этого синапса всегда равен 0 и электрический синапс всегда активен.
Мне это трудно принять Есть эфапс.
Нейроны
Описание формальных нейронов.
Сенсорный нейрон. Сенсорный нейрон – это нейрон с одним входом и одним выходом. При появлении сигнала на входе, он генерирует импульс с частотой, пропорционально амплитуде входного сигнала.
Почему с одним????? Сколько угодно. Вспомните ту же сетчатку.
(это просто биологическое дублирование J. Но если серьезно, то Вы правы, поэтому данное определение должно звучать так:
Сенсорный нейрон. Сенсорный нейрон – это нейрон с одним входом и множеством выходов. При появлении сигнала на входе, он генерирует импульс с частотой, пропорционально амплитуде входного сигнала.
Олаф )
Вставочный нейрон. Вставочный нейрон – это нейрон с двумя входами имеющий только электрические синапсы. Реализует функцию И по входу.
Очень узкое определение имеющее значение только в Вашей модели. Вставка может иметь любое число вх-вых (прим. автора: согласен, но другого названия для него не не нашлось, поэтому можете считать его вставочным нейроном Олафа J)
Ассоциативный нейрон. Ассоциативный нейрон имеет множество химических синапсов и один электрический. Мне каж это очень частный случай. Зачем электрический то??? Это особенности Вашей модели (прим. автора: прошу прощение за упрощение. Электрический в данной модели означает синапс, не имеющий веса, или имеющий бесконечный вес. Он может быть представлен как внесинаптическое гуморальное воздействие – описано ниже, так и простым электрическим синапсом. То есть имеется в виду, что ассоциативный нейрон имеет какое-то прямое воздействие на сому нейрона от какого-то фактора или на главный, самый толстый дендрит этого ассоциативного нейрона, а какое это воздействие, гуморальное или электрическое не важно. Лишь бы после этого воздействия произошла модификация химического синапса. В данной модели эл. синапс – это абстракция, но абстракция имеющая нейробиологический смысл) Это основной нейрон синаптической сети. При активации электрического синапса, во всех активных химических синапсах данного нейрона происходит модификация веса. Зачем во всех то – тогда какое же избирательное обучение может получиться. Но я об этом уже писал со ссылкой на статью (прим. автора: зачем во всех?! Для Вас я выделил жирным шрифтом – для активных в данный момент. Возможно, для Вас это будет уточнением, но для меня – это, само собой разумеется. Прошу прощения за …ну дальше вы знаетеJ)
Частота разряда нейрона прямо пропорциональна сумме весов всех активных синапсов этого нейрона. Не всегда. Сумматоры есть и внутри дендритов (примечание: в некоторых случаях электрический синапс может отсутствовать и сигнал на модификацию синапсов может приходить внесинаптическим путем) (прим. автора: активным синапсом считается синапс, у которого достаточно постсинаптических рецепторов на мембране для активации постсинаптического нейрона и само собой разумеется присутствие какого либо нейромедиатора внутри синаптической щели, а также других нейрогуморальных факторов, воздействующих непосредственно на нейрон. К сожалению это не полное определение активного синапса и автор это признает, но для той модели, что здесь описана, этого достаточно)
Промежуточные нейроны. Промежуточные нейроны имеют множественные электрические синапсы с аксонами ассоциативных нейронов. Частота разряда нейрона прямо пропорциональна сумме частот всех активных аксонов ассоциативных клеток имеющих электрические синапсы с этим нейроном.
Мне нравится, что Вы мыслите различными шаблонами отсеков, но это именно частные допущения Вашей модели
(прим. автора: Вадим, все эти «отсеки» появились лишь из простого предположения, что условный рефлекс можно реализовать не нанейронов, как это сделал один форумчанин из Томска, а всего на одном нейроне и двух синапсах. Ну, если быть более точным в Вашем понимании, то двух постсинаптических мембран на одном постсинаптическом нейроне и двух пресинаптических мембран на двух бутонах аксонов двух пресинаптических нейронах J )
Мотонейроны. Мотонейроны имеют множественные химические синапсы с аксонами промежуточных нейронов. Отличие от ассоциативных клеток в том, что у мотонейронов может быть активен только один синапс, вследствие торможения одним из промежуточных нейронов всех остальных промежуточных нейронов.
Мне каж что мотонейрон может быть таким же сумматором как и все остальные нейроны.
(прим. автора: а мне так не кажется. Неужели Вы думаете, что может быть один Мотонейрон Сумматор, который будет дифференцировнно выдавать сигнал на все мышечные нейроны? Мне кажется, что мотонейронов должно быть столько, сколько существует мышечных групп. Естественно, что существуют мыщцы антагонисты, и по этому естественно предположить, что на мотонейроне будут синапсы (или по Вашему аксон-дендритные комплексы J) как от нейронов одной так и от другой мышечной группы. И это вполне естественно, что сумма торможения и возбуждения на двух мотонейронах от двух мышц антагонистов будет соответствовать степени возбуждения и торможения этих двух мотонейронов, возбужденных разными синапсами от разных пресинаптических нейронов плюс обратная связь от силы сокращения каждой из мышц.
Правила
Изменение количества квантов медиатора в синапсе, при выработке условного рефлекса, происходит гетеросинаптически или моносинаптически, при одновременном внесинаптическом воздействии.??? (прим. автора: здесь имются ввиду гуморальные факторы - гормоны, прямое электрическое воздействие на нейрон и др. неучтенные или не открытые на данный момент факторы. Если Вы посмотрите современную литературу, то поймете о чем я говорил несколько лет назад.) Кстати, это очень принципиальный вопрос. Какой фактор заставляет синапс, или его мембраны изменять свою чувствительность и структуру.
Частота разряда пресинаптического нейрона модифицирует синапс пропорционально своей частоте. Нет. Есть еще фасилитация. То есть, чем выше частота, тем большее количество медиатора выбрасывается в синаптическую щель, и тем больший вес будет иметь данный синапс при выработке условного рефлекса. Нет. Может не быть соотв числа АМПА рецепторов – если пример для глутамата (прим. автора: исключения подтверждают правила…не я сказал J) (прим. автора: моделирование несколько отличается от копирования степенью упрощения не так ли?)
Частота разряда постсинаптического нейрона прямо пропорциональна сумме квантов медиатора во всех синаптических щелях этого нейрона. Нет. Медиатор бывает разный. И не только (прим. автора: а разве я говорю, что медиатор бывает один?!) Ясно, что появление медиатора в щели возможно только при прохождении сигнала через синапс. В неактивных синапсах медиатора в щели нет. Это так.
Аксиомы сети
1. Связи между нейронами в сети не случайны, а генетически детерминированы. Согласен.
В течении онтогенетического развития центральной нервной системы (ЦНС) некоторые синапсы создаются «не на том месте». Потом они рассасываются и заменяются новыми, правильно расположенными контактами. (Немечек, 1978)
Может это и слишком наивный взгляд нейробиологов. Генетика может оказаться не такой глупой. Я думаю, что распадаются управляющие связи, обеспечивающие вербовку нейронов при обучении а вовсе не неудачные связи J
(прим. автора: может быть…)
2. Пластичность жестко детерминированной сети обусловлена пластичностью
синапсов.
Не только синапсов
(прим. автора: а чем еще?)
Сами по себе пластические изменения синапсов можно рассматривать как способ хранения информации, который мог бы лежать в основе памяти. (Кендел, 1976)
Изменение синапсов это увеличение числа постинаптич. рецепторов и увеличение выброса медиатора или уменьшение естественно.
(прим. автора: ну да, это так…)
3. Условный рефлекс всегда дихотомичен. То есть он образуется всегда только между двумя сигналами. (Павлов, 1906, 1927), (Miller, Konorsky, 1928), (Skinner, 1938).
Оч. Сложный для меня вопрос. Обрабатывать входной сигнал двойками проще, но возможно что длина основания может быть большей. Тогда можно совмещать ассоциацию и логич. вывод в одной сети, а не в разных J
(прим. автора: это отдельный вопрос, требующий развернутой дискуссии, а пока оставим как есть.)
4. Каждая модальность имеет свой диапазон значений.
Не понял о чем это.
На этот вопрос коротко не ответишь. Ну, возьмем, к примеру, тонотопическую организацию дорсокаудальной (ДК) зоны слуховой области. 93% нейронов ДК-проекции обладают способностью избирательно отвечать генерацией ПД на предъявление звукового сигнала той или иной частоты. Обнаружена отчетливая зависимость между локализацией нейрона в данной проекционной зоне и значением его характеристической частоты (ХЧ). На каждом милиметре проекционной коры представлены звуковые частоты, охватывающие диапазон примерно 6.3 кГц. Установлена функциональная связь между пространственной локализацией нейроноы в ДК-зоне и значением его ХЧ. Чем дальше располагался нейрон в ДК-зоне от ее низкочастотного фокуса, тем выше было значение его ХЧ.
Данные, свидетельствующие о колонковой организации нейронов ДК-зоны: нейроны ДК-зоны, обнаруживаемые в различных слоях коры при вертикальном погружении электрода, имели достаточно близкие значения ХЧ. Такой тип организации нейронов ДК-зоны, по видимому, обусловлен наличием общих сенсорных каналов связи нейронов колонки с одним и тем рецептивным полем (РП) на базилярной мембране улитки. Эти данные согласуются с результатами, полученными при изучении функциональной организации нейронов слуховой области, зрительной и соматосенсорной областей коры в которых так же установлена колонковая организация относительно места и модальности раздражения. (, . Частотнотопическая организация дорсокаудальной зоны слуховой области AII коры мозга кошки//Нейрофизиология.-1988.-20, № 2.-с.220-227.)
5. Каждый диапазон значений имеет свой шаг. Например диапазон значений для слуха человека составляет от 20 догерц, а шаг диапазона составляет 3 герца. С меньшим шагом человек уже не различает изменение тона сигнала. То есть необходимо всего около 6000 сенсорных нейронов, расположенных в улитке слухового аппарата человека, чтобы перекрыть весь диапазон.
Ну это все особенности первичных рецепторов??
6. Количество сенсорных нейронов рассчитывается делением величины диапазона на его шаг.
Ну что то в этом есть. Другое дело – могут и есть перекрытия (прим. автора:Да, но мне кажется на более высоком уровне.)
7. Количество вставочных нейронов сети рассчитывается умножением количества сенсорных нейронов одной модальности на количество сенсорных нейронов другой модальности. То есть мы создаем избыточные потенциальные условно-рефлекторные связи между разными модальностями исходя из аксиомы 1. Так как все связи между нейронами закладываются в период формирования мозга и затем не меняются, а только модифицируются. Здесь необходимо акцентировать внимание на слове потенциальные, то есть в обученной сети их может быть на порядки меньше. Они образуются по мере необходимости.
Это может быть, мы это применяли несколько лет назад, но я перешел к более интересным архитектурам J J))) Язык показать?:))))
8. При проектировании разных сетей, архитектура сети не меняется, меняется только количество нейронов и синапсов. В природе принципы работы нейронных сетей одинаковы и у насекомых и у человека. По общему характеру активности нейроны гусениц не отличаются от нейронов позвоночных (Adrian, 1930). Он сравнил суммарную электрическую активность оптического ганглия (скопления зрительных нейронов) водного жука Dytiscus со своей собственной электроэнцефалограммой, отводимой от зрительной коры, и обнаружил сравнимые реакции на свет (Kandel, 1976)
Над этим я тоже иногда размышляю.
9. Основной элемент сети – синапс. Нет. (прим. автора: в данной модели – да.)
10. Основа архитектуры сети – условный рефлекс. Нет. УР – это просто следствие пластичности. (прим. автора: в данной модели это основное понятие, и архитектура сети построена в соответствии с этим)

11. Количество синапсов на ассоциативном нейроне равно количеству вставочных нейронов. Это следствие аксиомы 7. N*(N-1)
12. Количество ассоциативных нейронов вычисляется по формуле: K= 2 , где N – это количество модальностей.
13. Промежуточный (выходной) нейрон в однослойной сети всегда один.
14. Для получения большего выходного диапазона значений добавляются дополнительные однотипные сети, имеющие общие сенсорные и вставочные нейроны. По одной на каждый дополнительный выходной нейрон.
15. Количество моторных нейронов зависит от количества «мышц» или исполнительных устройств.
Наши сети существенно различаются, в них нет Вашей «универсальности». Мне каж. на Вас еще влияют парадигмы «классических» сетей???
Условный рефлекс
(базовые положения)
Связи между различными сенсорными модальностями
Это очень интересный и важный вопрос. Какая архитектура этих связей? От ответа на этот вопрос зависит, сможем мы построить работоспособную нейросинаптическую сеть или нет. В настоящий момент используется парадигма «все со всеми». Да наплюньте Вы на то, что там у них используетсяJ)))) Недоумки, прошу прошения.
Но эта парадигма приводит к проклятию размерности. Почему же в мозге нет этого проклятия? Потому, что мозг дихотомичен, двумерен, несмотря на многотысячные связи на одном нейроне. Из чего можно сделать такой вывод? Из понятия условного рефлекса. Условный рефлекс всегда дихотомичен???????????. Всегда устанавливается связь только между двумя событиями. Не уверен.
Условный рефлекс всегда дихотомичен. Это вытекает из самого определения условного рефлекса, как функции между безусловным и условным раздражителем.
Возможно ли существование условного рефлекса между тремя безусловными раздражителями и одним условным? Да такое сочетание реализуется с помощью трех условных рефлексов, где в каждом из них присутствует один условный раздражитель и один из трех безусловных. Думаю это один из возможных случаев. Но ИМХО могут несколько оснований активировать мишень.
Возможно ли существование условного рефлекса между тремя условными раздражителями и одним безусловным раздражителем?
Да такое сочетание реализуется с помощью трех условных рефлексов, где в каждом из них присутствует один безусловный раздражитель и один из трех условных.
Теорема 1. При множестве условных раздражителей мы всегда имеем одну и только одну реакцию.
Вариант 1. (частный случай) пусть B=>A, тогда при обучении мы получим
U1+B=A
U2+B=A
U3+B=A
после обучения мы получим набор соотношений
U1=>A
U2=>A
U3=>A
Это возможно.
Где U – условный раздражитель, B – безусловный раздражитель, A – событие (реакция) на безусловный или (после формирования условного рефлекса) на условный раздражитель.
То есть на три разных условных раздражителя мы получим одну и ту же реакцию.
Теорема 2. При одном условном раздражителе возможно существование множества различных реакций.
Вариант 2. пусть B1=>A1, B2=>A2, B3=>A3
Тогда во время обучения имеем набор уравнений
U+B1=A1
U+B2=A2
U+B3=A3
Тогда после обучения мы получим набор отношений
U=>A1
U=>A2
U=>A3
То есть на один условный раздражитель мы получим одновременно три различные реакции.
Введем новые функции в набор из N-го количества условных рефлексов – функцию силы связи Sn между Un и An. (и функцию торможения Tn между Un и Un+1.)
Поведение уравнений при первом и втором вариантах не изменится.
Рассмотрим
Вариант 3. пусть дано A1, A2, A3, B1, B2, B3, U1, U2, U3 тогда
Во время обучения
U1+B1=A1
U2+B2=A2
U3+B3=A3
После обучения
U1![]() A1 если S1>S2
A1 если S1>S2![]() S1>S3
S1>S3
U2![]() A2 если S2>S1
A2 если S2>S1![]() S2>S3
S2>S3
U3![]() A3 если S3>S1
A3 если S3>S1![]() S3>S2
S3>S2
Здесь мы тоже имеем только одну из трех возможных реакций из-за функции торможения сильной связью слабых.
Это всё возможные случаи но далеко не все возможныеJ.
По-видимому, для того, чтобы образовался условный рефлекс, должны существовать потенциальные синаптические связи для его возникновения, т. к. новые нейроны не образуются, а только отмирают. Образуются образуются http://www. salk. edu/faculty/faculty_details. php? id=23
Salk News Releases
Но в данном случае это не относится к делу в момент запоминания. Они образуются когда заканчиваются нейроны резерва и тогда начинают участвовать.
Так же на этот вывод указывает наблюдение о быстрой обучаемости и почти мгновенном образовании условного рефлекса в стрессовой ситуации. Но ведь если предположить, что аксоны растут по направлению активного нейрона и устанавливают синаптическую связь с дендритами другого нейрона в процессе обучения, то скорость роста аксона явно меньше скорости образования условного рефлекса.
Меня когда то тоже мучили эти вопросыJ)
Все вышесказанное однозначно указывает на то, что аксон-дендритные связи закладываются в головном мозге с самого начала, а затем активируются в процессе формирования условных рефлексов. При необходимости идет тот самый неонейрогенез, который вызвал вопросы у посетителей нейросайнз. (прим. автора: сайт помогает иногда взглянуть на проблему многими глазами и оценить «модные» веяния или узнать, что старые заблуждения, по прошествии многих десятков лет все еще в силе…)
Ведь наша нейронная сеть не просто отражает пассивно внешний мир, она устанавливает каузальные связи между событиями этого мира. Ведь просто увидеть какой либо предмет и не связать его с чем-то, мы этого предмета не увидим. Он просто не отразится в нашем сознании. Для того чтобы он отразился, его необходимо связать с каким либо событием (настоящим или прошлым). Этим и занимается условный рефлекс. Ну да. Он образует условно-рефлекторную связь между двумя событиями (т. к. объект можно тоже рассматривать как событие). Условный рефлекс является фундаментальной базовой функцией нервной системы. Теперь необходимо рассмотреть вопрос о его биологической или точнее нейробиохимической сущности.
УР – просто частный случай ассоциации, так ведь? (прим. автора: наверное Вы правы, но в данной модели схема возникновения УР и ассоциаций одна и та же, просто они происходят на разных уровнях и безусловный стимул на этих уровнях может быть представлен любым обучающим стимулом от других отделов сети. Суть и архитектура при этом остается одна и та же.)
На заре становления нейробионики условный рефлекс представляли в виде нейронной сети (см. рисунок) содержащей много нейронов.

Нервная сеть, моделирующая выработку условного рефлекса. (Брайнес, 1968)
Могут быть разные архитектуры для УР и для чего угодно. (прим. автора: это только один из примеров…)
Но если исходить из предпосылки, что главный элемент нейронной сети не нейрон, а синапс, тогда условный рефлекс можно представить в виде одного нейрона и двух синапсов, один из которых отвечает за безусловный раздражитель, а другой за условный. ЭТО Классический треугольник Хэбба. Можно даже предположить, что синапс, отвечающий за безусловный раздражитель является электротоническим, а синапс отвечающий за условный раздражитель – химическим. Прошу прощения, но искусственное допущение. (прим. автора: ещё раз хочу обратить внимание, что в данной модели под эл. синапсом на обучаемом нейроне подразумевается любое обучающее воздействие, не имеющее веса, или имеющего очень большой вес. Здесь речь может идти, как о внесинаптическом подкреплении группы нейронов, так и о конкретном воздействии на один нейрон. Это упрощение необходимо для более выпуклого представления архитектуры сети, иначе мы за деревьями дендритов не увидим леса сети.)
Модификация количества трансмиттеров в синаптической щели которого является весом синапса и силой условного рефлекса. Чем больше вес синапса или суммы синапсов на одном нейроне, тем с большей частотой передается сигнал по аксону.
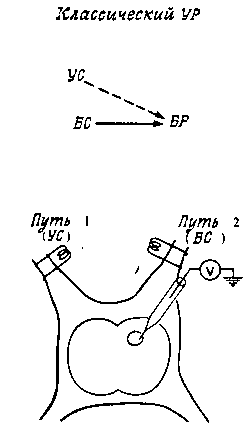
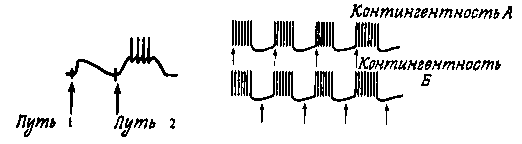
Схема опытов по выработке условного рефлекса на отдельном нейроне.(Kandel, 1967.)
Кендел подтвердил догадги Хэбба и описал каскад молекулярных изменений за ним стоящий за то и премию дали.
При распознавании образа состоящего из разных модальностей (слуха, зрения, вкуса и т. д. ) мы имеем составной условный рефлекс состоящий из пар: слух![]() зрение+B=A, слух
зрение+B=A, слух![]() вкус+B=A, зрение
вкус+B=A, зрение![]() вкус+B=A, где составной условный раздражитель выступает в роли одного условного раздражителя, а B выступает в роли одного безусловного раздражителя или внесинаптического химического подкрепления. НУ почему же внесинаптического то L((( ??? (прим. автора: см. выше…)Тогда составной условный рефлекс выражается отношениями Варианта 2(частный случай). Если же существует много событий А, тогда Вариант 2 и Вариант 3 объединяются в одну систему отношений:
вкус+B=A, где составной условный раздражитель выступает в роли одного условного раздражителя, а B выступает в роли одного безусловного раздражителя или внесинаптического химического подкрепления. НУ почему же внесинаптического то L((( ??? (прим. автора: см. выше…)Тогда составной условный рефлекс выражается отношениями Варианта 2(частный случай). Если же существует много событий А, тогда Вариант 2 и Вариант 3 объединяются в одну систему отношений:
U1+B1=A1
U2+B1=A1
U3+B1=A1
U1+B2=A2
U2+B2=A2
U3+B2=A2
U1+B3=A3
U2+B3=A3
U3+B3=A3
Ну это все семантические треугольники. Это все заивист от параметров отсеков.
Могут быть варианты, как расставишь коэффициенты (я имею в виду «наши» коэффициенты разных параметров отсеков, а не весаJ).
Может быть и
U1+B2=A2
и
U1+B2=A1
U1+B2=0
U1+B2=А1+А2
После обучения
U1![]()
U2![]()
![]()
![]() A1, если Sa1> Sa2
A1, если Sa1> Sa2![]() Sa1> Sa3
Sa1> Sa3
U3![]()
U1![]()
U2![]()
![]()
![]() A2, если Sa2> Sa1
A2, если Sa2> Sa1![]() Sa2> Sa3
Sa2> Sa3
U3![]()
U1![]()
U2![]()
![]()
![]() A3, если Sa3> Sa1
A3, если Sa3> Sa1![]() Sa3> Sa2
Sa3> Sa2
U3![]()
J)) как я не люблю формулы J))
Расчет времени задержки между условным стимулом и безусловнным
Для того, чтобы разобраться в этом, нам необходимо сначала разобраться в биологическом смысле этой задержке. Зачем она необходима. Многие исследователи просто констатируют, что для того, чтобы выработался условный рефлекс необходимо подавать безусловный стимул после условного с задержкой.
По всей видимости, эта задержка имеет глубокий биологический смысл.
J)) конечно – остаточные ВПСП в дендритах – вот и весь смысл J))))
Так условный рефлекс на вид открытого огня должен срабатывать до того, как рука попадет в зону его действия, вид и запах пищи должен повышать слюноотделение до того, как пища попала в рот, выделение желудочного сока должно происходить до того, как пища попала в желудок…
Из этих примеров видно, что время задержки должно быть разным. В некоторых случаях (не в эксперименте, а в нормальных условиях) она достигает нескольких часов. Так крысы, в естественных условиях, никогда не съедают отравленную приманку целиком, а откусывают маленькими порциями и ждут. Если через некоторый промежуток времени от 30 мин до нескольких часов не наступает симптомов отравления, то они откусывают еще и так далее. То есть время задержки между видом и запахом пищи и реакцией отравления довольно большое и все равно приводит к рефлексу избегания отравленной пищи. (Меннинг, 1979)
Да опыт это всеJ) Результат обучения.
Поэтому при проектировании синаптических сетей на основе условного рефлекса мы должны быть готовы к тому, что нам придется, исходя из задачи, вводить функцию задержки подкрепления условного рефлекса. Нет, самостоятельное значение задержки Вы преувеличиваете. J

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
