

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
2.4. Горох посевной.
Применение 9-АМА позволило впервые изучить рисунки OR - и С-окраски хромосом 8 сортов гороха посевного (Pisum sativum L.), который имеет размеры хромосом, сходные с ромашкой. При С-окрашивании хромосом гороха выявляются небольшие прицентромерные и теломерные, а также мелкие интеркалярные С-бэнды. На основании оценки морфологии хромосом и рисунков С-бэндинга была проведена идентификация хромосом в геноме гороха в соответствии со стандартной цитогенетической номенклатурой Бликста (Рис. 21а) [Blixt, 1959].
 |
Рис. 21. Кариотипы P. sativum L.: C- (а) и OR- (б) дифференциальное окрашивание хромосом. Компьютерное совмещение последовательно окрашенных хромосом ацетоорсеином (красный) и DAPI (синий) (в). 1-7 – номера хромосом в соответствии со стандартной номенклатурой Бликста.
Рисунки С-бэндинга хромосом у сортов овощного, сахарного, кормового и зернового гороха различались по размерам прицентромерных и приспутничных гетерохроматических блоков, а также интеркалярных С-бэндов в коротких плечах 2, 3, 4 и 7 хромосом и в длинных плечах хромосом 1 и 5.
Четкий OR-бэндинг c большим числом темноокрашенных районов был выявлен на хромосомах тех же сортов гороха. По рисункам OR-окраски идентифицированы все хромосомы (Рис. 21б). Также как и С-окрашенные хромосомы, они были расположены по размерам и центромерному индексу в соответствии с номенклатурой Бликста. Было проведено последовательное (Рис. 21в) OR-, а затем С/DAPI-окрашивание (Рис. 21а и 22а) хромосом гороха, которое подтвердило правильность такого подхода при их идентификации. Межсортовых различий по рисункам OR-окраски не было выявлено.
При исследовании хромосом гороха было обнаружено, что на профазных и прометафазных хромосомах после предобработки 9-АМА и процедуры FISH выявляется Q-подобный DAPI-бэндинг. В зависимости от фазы митоза DAPI-бэндинг может быть различной степени разрешения. По мере компактизации хромосом Q-подобный DAPI-бэндинг переходит в С-подобный DAPI-бэндинг (C/DAPI-бэндинг) (Рис. 22а).
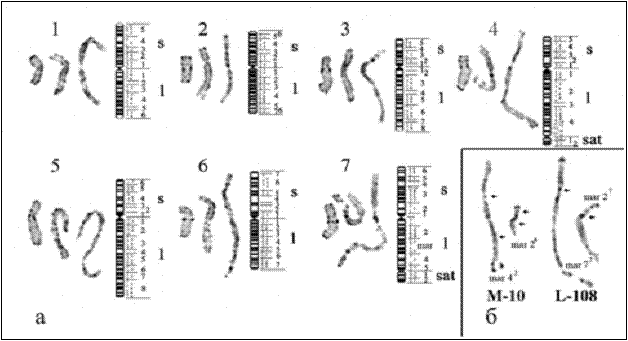
Рис. 22. Кариограммы и идиограммы DAPI-окрашенных (инвертированное изображение) хромосом гороха (1-7 – номера хромосом в соответствии со стандартной номенклатурой Бликста): а – разные степени разрешения DAPI-бэндинга (на профазных хромосомах суммарное число DAPI-полос на гаплоидный набор хромосом гороха равно 344); б – точная локализация точек разрывов хромосом в транслокационных линиях гороха L-108 и M-10 по рисункам высокоразрешающего DAPI-окрашивания.
Это обстоятельство позволяет подобрать степень конденсации хромосом, при которой по рисунку DAPI-бэндинга возможно идентифицировать хромосомы и при необходимости локализовать на них точки хромосомных перестроек или ДНК-зонды. С помощью такого подхода была изучена структура кариотипов транслокационных линий L-108 и M-10 гороха. DAPI-бэндинг высокого разрешения позволил точно установить на профазных-прометафазных хромосомах точки разрыва хромосом при транслокациях (Рис. 22б). Для линии L-108 выявлены точки разрывов в проксимальных районах коротких плеч хромосом 2 и 7. Для линии М-10 впервые установлено наличие двойной транслокации между длинными и короткими плечами хромосом 2 и 4.
Сравнительное исследование рисунков DAPI-бэндинга хромосом у всех восьми сортов и 2 родственных линий (с высоким симбиотическим потенциалом) гороха, выявленных после FISH картирования рибосомных генов и теломерного повтора, обнаружило невысокий уровень внутри - и межсортового полиморфизма, подобно С-окраске (Рис. 23).
У всех изученных образцов гороха 26S рДНК находится в районах вторичных перетяжек и на спутнике 4 хромосомы и в приспутничном районе 7 хромосомы. Сайты 5S рДНК располагались на 1, 3 и 5 хромосомах, причем высокоразрешающее DAPI-окрашивание дало возможность уточнить районы локализации 5S рДНК. Теломерные повторы выявлялись на концах всех хромосом (см. раздел 1.3.1, рис. 6) как в нормальных, так и в транслокационных кариотипах. Дополнительных сайтов гибридизации с рДНК и теломерными последовательностями не выявлено (Рис. 23).
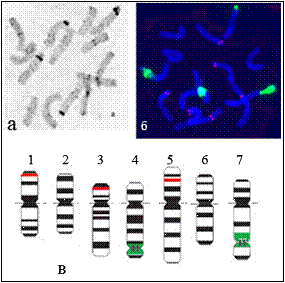 Ag-ЯОР-окрашивание показало, что в изученных образцах гороха оба локуса 26S рДНК, расположенные на хромосомах 4 и 7, функционально активны. В линиях гороха, несущих транслокации по ЯОР-образующим хромосомам 4 и 7, изменений функциональной активности ЯОР в этих хромосомах не обнаружено.
Ag-ЯОР-окрашивание показало, что в изученных образцах гороха оба локуса 26S рДНК, расположенные на хромосомах 4 и 7, функционально активны. В линиях гороха, несущих транслокации по ЯОР-образующим хромосомам 4 и 7, изменений функциональной активности ЯОР в этих хромосомах не обнаружено.
Рис. 23. Хромосомы гороха сорта Frisson: а - С-бэндинг, б - FISH-картирование 45S (зеленый) и 5S (красный) рДНК, в - идиограмма хромосом с указанием расположения С/DAPI-бэндов (черный), 45S (зеленый) и 5S (красный) рДНК 1-7 – номера хромосом в соответствии со стандартной номенклатурой [Blixt, 1959; Fuchs et al., 1998].
Исследование геномного полиморфизма у изученных сортов и линий гороха методом RAPD-PCR-анализа в полиакриламидном геле с 3 информативными праймерами А06, А09 и А10 в выявило между ними различия по 45 полиморфным локусам. RAPD-спектры всех исследованных представителей сорта или линии делились на кластеры в четком соответствии с сортовой принадлежностью. В кластерах линии Sprint 2 и сорта Finale обнаружен геномный полиморфизм между изученными индивидуальными растениями. Полученные нами результаты подтверждают возможность применения RAPD-анализа для идентификации сортов гороха [Гостимский и др., 2005].
Таким образом, значительных кариотипических различий исследованных сортов и линий гороха выявлено не было. Тем не менее, даже небольшие специфические особенности их геномов могут служить основой для создания кариогеномного паспорта. Такой подход может быть особенно важен для сертификации определенного генотипа гороха посевного при создании новых сортов и проверке чистоты суперэлитного и элитного семенного материала.
2.5. Исследование Zingeria biebersteiniana и родственных видов.
2.5.1. Цингерия Биберштейна.
Наряду с исследованием мелкохромосомных видов нами были изучены дикорастущие злаки, с числом хромосом кратным 2 – Z. biebersteiniana (Claus) P. Smirn. (2n = 4) и Z. pisidica (Boiss.) Tutin (2n = 8), Colpodium versicolor (Stev.) Schmalh. (2n = 4) с целью изучения формирования и происхождения их уникальных двухромосомных геномов. Сложность исследования хромосом этих растений была связана с тем, что, несмотря на довольно крупные размеры хромосом (8-11 мкм), две пары хромосом цингерии не всегда легко идентифицируются по размерам и морфологии, особенно в случае отсутствия выраженных вторичных перетяжек, а рисунок С-бэндинга у них беден и поэтому малоинформативен. При С-окрашивании хромосом Zingeria biebersteiniana четко выявляются только крупные С-бэнды в прицентромерных районах хромосом, а интеркалярные и прителомерные С-бэнды имеют очень маленькие размеры и выявляются нерегулярно (Рис. 23а). Следует отметить, что по рисункам С-бэндинга идентификация хромосом цингерии не составляет труда, но сравнение хромосом близкородственных видов значительно затруднено ввиду плохой выраженности интеркалярных С-бэндов. Это обстоятельство выдвинуло на первый план задачу поиска дополнительных хромосомных маркеров, позволяющих детально исследовать структуру хромосом двухромосомных злаков.
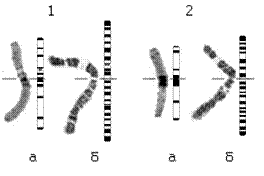 Применение 9-АМА позволило выявить на недоконденсированных хромосомах длиной 11-15 мкм чёткий рисунок OR-бэндинга (Рис. 23б).
Применение 9-АМА позволило выявить на недоконденсированных хромосомах длиной 11-15 мкм чёткий рисунок OR-бэндинга (Рис. 23б).
Рис. 24. Кариограммы и идиограммы цингерии: а - C- бэндинг и б - OR- бэндинг. 1 и 2 – номера хромосом.
По сравнению с С-окраской рисунок OR-бэндинга в хромосомах цингериии представлен большим числом интеркалярных бэндов. При этом С-позитивные прицентромерные районы хромосом, а также области вторичных перетяжек ацеторсеином не прокрашиваются.
2.5.2. Происхождение двухромосомных злаков Zingeria и Colpodium.
Для установления путей происхождения уникальных двухромосомных геномов злаков проведено сравнительное исследование хромосом в кариотипах Z.biebersteiniana (2n = 4), Z. pisidica (2n = 2х = 8), Colpodium versicolor (2n = 4) и Catabrosella variegata (syn.= Colpodium variegatum) (2n = 10). Метод С-дифференциального окрашивания выявляет на хромосомах этих видов малоинформативные рисунки, тем не менее, в кариотипах двух образцов Z. bibersteiniana из разных мест произрастания обнаружен межпопуляционный полиморфизм С-блоков хромосом. В частности, хромосомы растений, собранных вблизи лимана Пришиб в Волгоградской области, содержат более крупные прителомерные и интеркалярные С-блоки по сравнению с хромосомами растений, выросших в окрестностях с. Рахинка той же области.
 |
Рис. 25. Кариограммы, идиограммы, метафазные пластинки С-окрашенных хромосом и FISH с зондами 45S (зеленый) и 5S (красный) рДНК. Zb –Z.bibershteniana, Zp – Z.pisidica и C – C.versicolor. Масштабные линейки по 5 мкм.
Сравнительный анализ геномов C. versicolor и Z. biebersteiniana показал, что они сходны по морфологии и характеру распределения интеркалярных С-блоков по одной из хромосом, тогда как по другой (ядрышкообразующей) хромосоме различаются по центромерному индексу, положению вторичных перетяжек и рисунку С-окрашивания.
Установлено, что две пары хромосом из четырех в кариотипе Z. pisidica имеют значительное сходство с хромосомами Z. biebersteiniana. Однако, в отличие от Z. biebersteiniana, хромосомы Z. pisidica не содержат крупных прицентромерных С-блоков. Две другие хромосомы по рисункам С-окраски сходны с хромосомами C. versicolor. Однако размеры отдельных интеркалярных С-блоков у Z. pisidica значительно крупнее, чем таковые в хромосомах C. versicolor (Рис. 25). По-видимому, один из субгеномов Z. pisidica и геном Z. biebersteiniana имеют общее происхождение. Не исключено, что предковый геном C. versicolor принимал участие в формировании другого субгенома Z. pisidica.
У исследованных видов проведена хромосомная локализация генов 45S и 5S рРНК. Рисунки расположения генов 45S и 5S рРНК на хромосомах двух видов цингерий и у C. versicolor подтверждают предположение, что Z. pisidica является аллотетраплоидом, у которого один из субгеномов сходен с геномом Z. biebersteiniana (Рис. 24). Сравнение кариотипов Catabrosella variegata и описанных выше видов злаков не выявило черт сходства между ними.
Проведено сравнение последовательностей внутренних транскрибируемых спейсеров ITS1 и ITS2 генов 45S рРНК в геномах изученных видов. Обнаружено, что геномы уникальных двухромосомных злаков (x = 2) – Z. biebersteiniana (2n = 4), Z. pisidica (2n = 2х = 8) и Colpodium versicolor (2n = 4), относимые ранее к разным трибам или подтрибам, представляют два близких рода, генетическое расстояние (p-distance) между ITS которых составляет всего 1,2–4.4%. На молекулярно-филогенетическом древе виды Zingeria и Colpodium versicolor образуют единую кладу с Catabrosella araratica (2n = 42, x = 7).
Таким образом, установлено, что геномы Zingeria и Colpodium с наиболее низким из известных основным числом хромосом (x = 2) возникли не в разных филогенетических ветвях, а имеют монофилетическое происхождение.
2.6. Изучение геномов у видов рода Linum L.
Для определения родственных взаимоотношений и уточнения таксономического статуса, а также выявления филогенетических взаимосвязей видов рода Linum L. проведено изучение геномов возделываемого Linum usitatissimum L. и 25 дикорастущих видов из 6 секций [Юзепчук, 1949; Егорова, 1996], в основном произрастающих в Евразии.
2.6.1. Идентификация хромосом, геномы и кариотипы видов льна.
Род Linum – лен включает около 200 видов, которые разделяют на 5-6 секций [Юзепчук, 1949; Ockendon, Walters, 1968; Егорова, 1996]. Кариологическое изучение видов льна начато более полувека назад, и почти у 30 видов Нового и 50 видов Старого Света были определены хромосомные числа. Показано, что в кариотипах льновых число хромосом варьирует от 12 до 84. Однако, у многих видов льна числа хромосом, определенные разными исследователями, различались. Даже в кариотипе наиболее хозяйственно важного возделываемого вида L. usitatissimum L. насчитывали два числа хромосом 2n = 30 и 2n = 32. Хромосомы у льнов имеют сходную морфологию (в основном это метацентрики), и их размеры колеблются от 1 мкм до 6 мкм [Ray, 1944; Болховских и др.,, 1969; Rogers и др., 1972; Chennaveeraiah, Joshi, 1983].
При изучении кариотипов видов из 6 секций рода Linum L. нами впервые установлены хромосомные числа у L. squamulosum Rudolphi (2n = 18), L. komarovii Juz. (2n = 18), L. thracicum Degen. (2n = 28+1-6B) и L. czernjajevii Klokov (2n = 28+1-6B); уточнены хромосомные числа у L. decumbens Desf. ( 2n = 16), L. narbonense L. (2n = 28), L. stelleroides Planch. (2n = 18), L. pallescens Bunge (2n = 18). На основании исследований кариотипов 22 долгунцовых и масличных сортов льна, а также межеумков было подтверждено, что число хромосом у L. usitatissimum – 2n = 30.
До настоящего исследования ни у одного вида льна по результатам монохромного окрашивания в кариотипе не были идентифицированы пары гомологов. В результате дифференциального окрашивания в хромосомах всех изученных видов были выявлены С/DAPI-бэнды, самые крупные из которых располагались преимущественно в прицентромерных районах, а средние и небольшие – в теломерных и интеркалярных районах хромосом. На хромосомах всех изученных видов были локализованы пробы 26S и 5S рДНК. В геномах большинства видов льнов обнаружены 1-2 основных локуса 26S рДНК, ко-локализованные с 5S рДНК, а также 1-3 отдельных локуса 5S рДНК. Исключение составили два вида секции Linopsis (syn.= Linastrum), в геномах которых 5 из 9 пар хромосом несут сайт 26S рДНК и одна – 5S рДНК. На основании изучения морфологии хромосом, рисунков С/DAPI-окрашивания и результатов FISH-анализа удалось провести полную идентификацию хромосом в кариотипах всех исследованных видов. В кариотипах видов секции Syllinum впервые были выявлены и охарактеризованы В-хромосомы.
2.6.2. В-хромосомы.
В кариотипах видов секции Syllinum: L. flavum L., L. capitatum Kit. еx Schultes, L. campanulatum L., L. thracicum Degen, L. tauricum Willd, L. elegans Sprun. ex Boiss., L. czerniajevii Klokov нами были обнаружены добавочные хромосомы (В-хромосомы), число которых варьировало в клетках корневой меристемы от 1 до 6, а у некоторых растений эти хромосомы отсутствовали вовсе.
Установлено, что В-хромосомы в кариотипах исследованных видов имеют сходную структуру. В подавляющем большинстве случаев они представляют собой субметацентрики длиной около 1 мкм, и являются самыми маленькими хромосомами в наборе. Они гетеропикнотичны, интенсивно окрашиваются DAPI и СМА (Рис. 26-III), содержат множественные сайты 5S и 26S рДНК (Рис. 26-IV), которые не проявляют функциональной активности (Рис. 26-II). В - хромосомы с такой морфологией мы назвали "основным 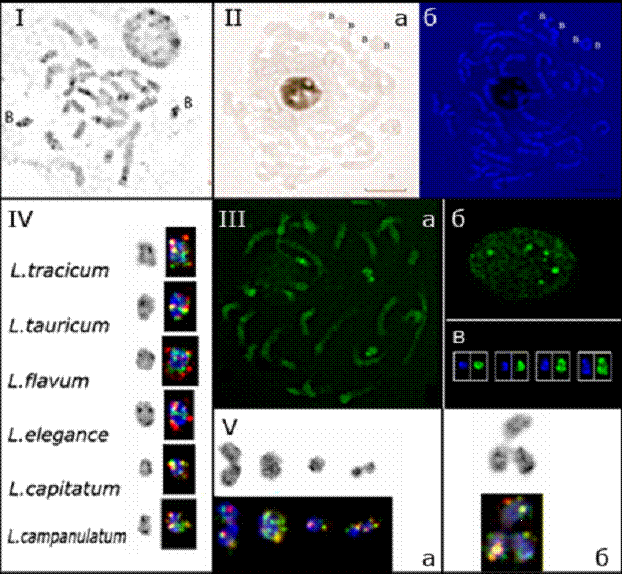
морфологическим типом" (Рис. 26-IV).
Рис 26. В-хромосомы видов секции Syllinum. I – С-окраска хромосом и ядра L.flavum; II – Ag-ЯОР-окрашивание профазных хромосом L.flavum (а) и его совмещение с последующим DAPI-окрашиванием (б); III – CMA-окрашивание В-хромосом в профазе (а), ядре (б) и сравнение рисунков окраски CMA и DAPI метафазных В-хромосом; IV – основной морфологический тип В-хромосом у разных видов, представлено инвертированное изображение DAPI-окрашивания и рядом та же хромосома с локализацией 26S рДНК (зеленый сигнал) и 5S рДНК (красный сигнал); V – иногда встречаемые атипичные В-хромосомы (а) и ассоциации В-хромосом (б).
Иногда встречаются другие морфологические типы В-хромосом – метацентрики и мелкие акроцентрики (Рис. 26-Vа). Возможно, что такие "атипичные" хромосомы образуются из В-хромосом основного типа в результате хромосомных перестроек. В некоторых метафазных пластинках обнаружены В-хромосомы, объединённые в кластеры, напоминающие по форме грозди винограда (Рис. 26-Vб). Следует отметить, что хотя по размеру и рисунку DAPI-бэндинга В-хромосомы иногда бывает трудно отличить от самых мелких хромосом постоянного набора (А-хромосом), наличие генов рибосомных РНК в В-хромосомах позволяет надежно их идентифицировать. Исследование В-хромосом у видов секции Syllinum позволило точно установить число А-хромосом в их кариотипах (2n = 28).
2.6.3. Сравнительные молекулярно-цитогенетические исследования видов льна.
Все исследованные виды льнов были разделены на 8 групп на основании сходства их кариотипов. Виды, объединенные в одну группу, имели сходную структуру кариотипов, а межвидовые различия, выявленные по рисунку С/DAPI-окраски хромосом и расположению рибосомных генов, были представлены небольшим числом реципрокных хромосомных транслокаций или перицентрических инверсий. В то же время, кариотипы видов, входящих в разные группы, значительно отличались друг от друга как по размерам, числу хромосом и рисункам их дифференциального окрашивания, так и по числу и расположению на хромосомах рДНК локусов. Кариограммы и идиограммы одного из видов, принадлежащих к каждой группе представлены на рисунке 27.
Первую группу составили виды с 2n = 18 из секций Adenolinum и Stellerolinum. Хромосомы вида L. stelleroides Planch., который систематики выделяют в монотипную секцию Stellerolinum Juz. ex Probat. [Юзепчук, 1949], по рисунку С/DAPI-окраски и расположению рибосомных генов оказались схожими с видами из секции Adenolinum (L. austriacum subsp. austriacum L., L. austriacum subsp. еuxinum Juz. (=L. squamulosum Rudolphi), L. leonii F. W. Schultz, L. perenne L., L. lewisii Pursh., L. komarovii Juz., L. altaicum Ledeb., L. mesostylum Juz., L. pallescens Bunge.). По-видимому, морфологические и анатомические отличия видов этих секций обусловлены различиями их геномов на молекулярном уровне, не детектируемыми цитогенетическими методами.
Секция Linum разделилась три группы с 2n = 30, 2n = 16 и 2n = 28. Следует отметить, что близкородственные 30-хромосомные виды L. usitatissimum L. и L. bienne Mill. отличаются от L. angustifolium Hunds. перицентрической инверсией хромосомы 3. Эти виды и виды L. grandiflorum Desf. и L. decumbence Desf. с 2n = 16 секции Linum, хотя и различаются плоидностью, но похожи по рисункам С/DAPI-окраски многих хромосом набора и расположению рибосомных генов. Высокое сходство рисунков С/DAPI-бэндинга наблюдается также между этими видами секции Linum и видами секций Adenolinum и Stellerolinum. При этом кариотипы последних, по нашим данным, отличаются от 16-хромосомных видов по наличию, по крайне мере, одной транслокации и дупликации по хромосоме 8. Таким образом, результаты хромосомного анализа подтверждают общность происхождения вышеуказанных видов секций Linum, Adenolinum и Stellerolinum.

Рис. 27. Кариограммы и идиограммы DAPI/С-окрашенных хромосом с локализацией 26S и 5S рДНК представителей различных секций рода Linum L. * –Знаком (?) помечены спорные таксоны.
Вид L. narbonense L. резко отличался как по числу и размерам хромосом, так и по рисунку их С/DAPI-окраски и расположению рибосомных генов от других видов секции Linum. Анализ хромосом по молекулярно-цитогенетическим маркерам выявил тетраплоидную природу генома этого вида. По морфологии и размерам хромосомы L. narbonense скорее схожи с таковыми у L. hirsutum L. секции Dasylinum и видами секции Syllinum (2n = 28+B), что согласуется с мнением ботаников, считающих L. narbonense переходным видом, систематическое положение которого нуждается в уточнении [Светлова, 2007; Оптасюк, 2007].
Как уже говорилось выше, обнаружение в кариотипах видов L. flavum, L. capitatum, L. campanulatum, L. thracicum, L. tauricum, L. elegans, L. czerniajevii секции Syllinum В-хромосом, позволило установить, что число хромосом в основном хромосомном наборе этих видов равно 28. Обнаружено, что основные А-хромосомные наборы всех этих видов сходны по размерам хромосом, рисунку C/DAPI бэндинга и локализации рДНК. У двух видов L. capitatum и L. czerniajevii выявлено по одной реципрокной транслокации хромосом.
Особняком от остальных видов секции Syllinum стоит 26 хромосомный вид L. nodiflorum L. Хромосомы этого вида имеют значительно меньшие размеры по сравнению с хромосомами других видов этой секции, В-хромосом у этого вида не обнаружено. Кроме того, L. nodiflorum отличается от других видов секции по расположению и количеству локусов генов рибосомных РНК. Следует, однако, отметить, что, несмотря на различия в размерах, хромосомы L. nodiflorum схожи с хромосомами других видов секции по рисунку С/DAPI-бэндинга. Последнее указывает на то, что L. nodiflorum имеет все же отдаленное родство с 28-хромосомными видами секции. Результаты молекулярно-цитогенетического анализа позволяют предположить, что в процессе формирования кариотипа L. nodiflorum имело место по крайней мере одно хромосомное слияние. Выявленные геномные отличия L. nodiflorum от других видов секции Syllinum хорошо согласуются с мнением некоторых систематиков о необходимости выделения этого вида в отдельную секцию Tubilinum [Светлова, 2006].
Две отдельные группы составляли виды секций Dasilinum и Linopsis. Кариотип L. hirsutum L. subsp hirsutum секции Dasylinum состоит из 16 хромосом, имеющих самые крупные размеры среди льновых. По рисункам С/DAPI-бэндинга и расположению рибосомных генов хромосомы этого вида схожи с отдельными хромосомами L. narbonense, L. nodiflorum и видов из секции Linopsis.
Кариотипы L. suffruticosum L. subsp. salsoloides (Lam.) Rouy. и L. tenuifolium L. секции Linopsis (syn. Linastrum) представлены 9 парами сходных по рисункам С/DAPI-бэндинга хромосом. Эти виды различаются по наличию транслокации между хромосомами, несущими гены рРНК. Поэтому у L. tenuifolium на одной из 5 спутничных хромосом в разных плечах расположены сайт ко-локализованных 26S и 5S рДНК и отдельный сайт 5S рДНК. Таким образом, у этих видов представлены практически все морфологические типы хромосом, несущих гены рРНК, которые имеются у видов из других секций.
Параллельно с комплексным хромосомным анализом для этих же видов льна был проведен анализ геномного полиморфизма с помощью высокоразрешающнго метода RAPD-PCR в полиакриламидном геле. В результате была построена UPGMA-дендрограмма геномной близости видов рода Linum (Рис. 28).
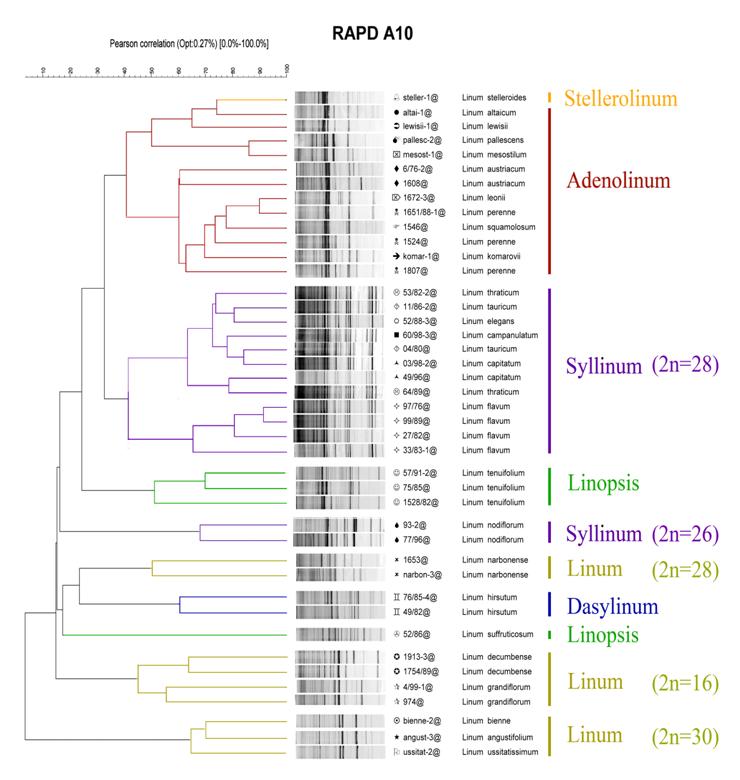
Рис. 28. Дендрограмма генетической близости видов различных секций рода Linum L., построенная по результатам RAPD-анализа.
Результаты RAPD-анализа хорошо согласуются с данными кариологического исследования. Обнаружено, что группы видов, имеющие сходную структуру кариотипа, имеют и низкий уровень межгеномной дивергенции и кластеризуются совместно. Так, виды секций Adenolinum и Stellerolinum попали в один кластер, а L nodiflorum кластеризуется отдельно от 28-хромосомных видов секции Syllinum. Отдельный кластер формирует вид секции Dasylinum. 30- и 16- хромосомные виды Linum и L. narbonense также формируют три отдельных кластера. Исключение составили виды секции Linopsis: L. suffruticosum subsp. salsoloides (Lam.) Rouy. и L. tenuifolium L. Для них выявлен высокий уровень межгеномной дивергенции, несмотря на то, что, по нашим данным, их кариотипы различаются по одной реципрокной транслокации. Систематика этих видов до сих пор не вполне ясна. У них была выявлена значительная межпопуляционная изменчивость по морфологическим признакам, и в то же время существует мнение, что L. suffruticosum и L. tenuifolium являются подвидами, а не самостоятельными видами [Nicholls, 1985].
2.6.4. Хромосомные числа видов рода Linum.
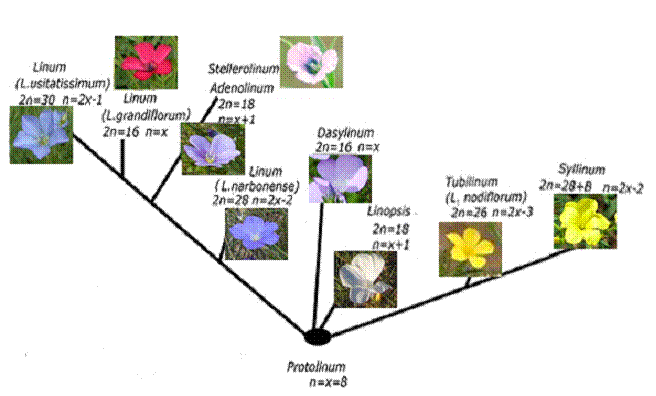
Предполагается, что предковая форма рода Linum (Protolinum) могла иметь основное число хромосом равное 8 или 9. Эти числа встречаются у диплоидных видов рода, отнесенных к разным филогенетическим ветвям. Виды секций Adenolinum, Stellerolinum и Linopsis имеют n = 9, а виды секции Linum, Dasylinum и Cathartolinum имеют n = 8.
Рис. 29. Гипотетическое эволюционное древо хромосомных чисел рода Linum L.
Ранее на основании анализа кариотипов при монохромном окрашивании хромосом было построено предполагаемое эволюционное древо рода Linum, в основание которого был помещен диплоидный вид c основным числом хромосом n = x = 9 [Chennaveeraiah and Joshi, 1983]. Если предположить, что исходным было число хромосом n = x = 8, то оказывается, что редукция хромосомных чисел имела место только у полиплоидных видов льна (n = 13, 14, 15), а диплоидные виды с n = 9, 10 могли возникнуть вследствие хромосомных дупликаций. На наш взгляд это наиболее простое объяснение изменчивости основного числа хромосом у льнов (Рис. 29).
Как показало проведенное нами кариологическое исследование, в процессе видообразования льновых происходила полиплоидизация, различные хромосомные перестройки, в частности, хромосомные слияния, а также могла иметь место полисомия и утрата хромосом полиплоидами, что и привело к существующему разнообразию хромосомных чисел у видов рода Linum. Поэтому в настоящий момент не представляется возможным достоверно установить, какое именно основное хромосомное число было исходным у льнов. Возможно, что это число было даже более низким, чем у ныне живущих диплоидных видов. В связи с этим следует упомянуть, что на основании недавно проведенного молекулярно-филогенетического исследования последовательностей внутренних нетранскибируемых спейсеров генов 45S рРНК и нескольких генов генома хлоропластов, было высказано мнение, что род Linum не является монофилетическим [McDill et al., 2009]. Если это действительно так, то единого для рода Linum исходного хромосомного числа может и вовсе не существовать.
Таким образом, сравнительное молекулярно-кариологическое исследование видов рода Linum L. позволило подтвердить их филогенетические связи. В то же время обнаружено, что систематика рода не вполне совершенна, и хочется надеяться, что полученные нами результаты помогут уточнить таксономию рода Linum L.
Заключение.
В работе представлен комплекс методов, позволяющих осуществлять детальное изучение хромосомной организации геномов растений, включая надежную идентификацию малоинформативных (небольшие размеры и/или бедность рисунков С-окраски) индивидуальных хромосом. Использование метода выявления рисунков ранней репликации хромосом открывает широкие возможности не только для идентификации хромосом, но и для более глубокого исследования последовательности репликации ДНК у растений. Применение ДНК-интеркаляторов тормозит процесс конденсации митотических хромосом, что приводит к повышению разрешающей способности дифференциального окрашивания и FISH с различными маркерными последовательностями ДНК. Оптимизация программно-аппаратного комплекса регистрации и анализа изображений значительно облегчает, ускоряет и делает более объективными исследования хромосомной организации геномов растений даже с очень мелкими хромосомами. Четкая идентификация индивидуальных хромосом на препаратах, полученных с помощью щадящих структуру ДНК процедур, может служить в качестве кариологической основы при осуществлении проектов тотального секвенирования геномов хозяйственно-ценных мелкохромосомных растений, поскольку открывает возможность получать хромосомо - и локус-специфические клонотеки с помощью микродиссекции и способствовать созданию генных контигов различной протяженности.
Применение комплексного подхода для одновременного сравнительного исследования кариотипических особенностей и геномного полиморфизма видов льна показало, что на уровне секций результаты, полученные с помощью цитогенетических и молекулярных методов полностью совпадают. На видовом уровне крупные геномные реорганизации, такие как хромосомные перестройки, выявляются методами хромосомного анализа, а мелкие геномные реорганизации – методом RAPD-анализа. Таким образом, с помощью RAPD-анализа можно оценить степень генетической близости родственных геномов, а хромосомный анализ позволяет выявить различия в хромосомной организации геномов, и, следовательно, оба подхода хорошо дополняют, но не заменяют друг друга. Результаты исследования геномов видов секций Linum, Syllinum, Adenolinum, Stellerolinum, Dasylinum и Linopsis важны не только для решения разнообразных проблем эволюционной и сравнительной геномики льна, но и для развития генетических и молекулярных основ селекции льна посевного. Изучение геномов различных видов льновых продолжается, однако уже сейчас можно утверждать, что полученные нами сведения об их кариотипах заложили цитогенетические основы для проведения тотального секвенирования генома L. usitatissimum L. Осуществление этого проекта помогло бы разрешить многие проблемы селекции культурного льна, а также систематики и эволюции видов рода Linum L. на основе новых сведений, которые можно получить методами сравнительной геномики.
ВЫВОДЫ
1. Разработан комплекс цитогенетических методов (выявление рисунка ранней репликации хромосом растений, задержка конденсации митотических хромосом с помощью 9-АМА, OR-бэндинг, Q-подобный DAPI-бэндинг и применение новых флуоресцентных лигандов, созданных на базе красителя Hoecht, - DB(8) и DB(17), для получения высококонтрастного флуорохромного бэндинга), и повышающий разрешающую способность анализа хромосомной организации геномов растений с хромосомами малых размеров или малоинформативным рисунком дифференциального окрашивания.
2. Показано, что FISH-картирование универсальных молекулярных зондов (рДНК, теломерный повтор) не только облегчает идентификацию хромосом и выявление хромосомных перестроек в кариотипах растений с хромосомами малых размеров или малоинформативным рисунком С-бэндинга, но и дает возможность более точно устанавливать взаимотношения родственных геномов. Для растений с небольшими геномами и мелкими хромосомами FISH-картирование геномной ДНК или ее фрагментов из геномных библиотек перспективно для получения дополнительных маркеров, позволяющих проводить успешную идентификацию их хромосом.
3. Применение последовательно или одновременно нескольких молекулярно-цитогенетических методов в сочетании с оптимизацией программно-аппаратного комплекса получения и анализа изображений значительно расширяет возможности исследования хромосомной организации геномов растений с хромосомами малых размеров или малоинформативным рисунком дифференциального окрашивания.
4. По рисункам ранней репликации идентифицированы 13 пар гомологов в геноме A1 диплоидного хлопчатника G. herbaceum и 26 пар - в геноме (AD)2 аллотетраплоидного хлопчатника G. barbadense. Хромосомы генома (AD)2 были разделены на субгеномы Ab и Db. Сходство рисунков ранней репликации не только в A1 и Ab геномах, но и в Db геноме свидетельствует о консервативности последовательности репликации ДНК в родственных геномах.
5. С применением 9-АМА впервые проведено морфометрическое исследование хромосом одноклеточных красных водорослей. Показано, что число хромосом может быть использовано для уточнения таксономии класса Cyanidiophyceae (Rhodophyta). Обнаружено, что изученные виды, относящиеся к наиболее примитивным эукариотам, обладают и практически самыми малыми геномами, что демонстрирует зависимость хромосомной организации генома, от его размера.
6. Cравнительное исследование хромосом по рисункам С - и OR-бэндинга в кариотипах трех сортов ромашки аптечной ( M. chamomilla L.) показало, что они различались по полиморфизму С-блоков и не различались по рисункам OR-бэндинга. По сходству рисунков С-окраски хромосом обнаружено, что дикорастущий вид M. inodora является автотетраплоидом, субгеномы которого сходны с геномом M. maritima , который был обозначен Mm. Его сравнение с Mch-геномом ромашки аптечной выявило сходство морфологии хромосом и рисунков распределения позиций гетерохроматических районов по длине хромосом, что говорит об общности их происхождения.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
