

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
а
Хромосомная организация геномов растений
с хромосомами малых размеров или малоинформативным рисунком дифференциального окрашивания.
Специальности: 03.01.03 – молекулярная биология
03.01.15 - генетика
Диссертация в виде научного доклада
на соискание ученой степени
доктора биологических наук
Москва 2010
Работа выполнена в Лаборатории молекулярной кариологии и основ клеточной терапии Учреждения Российской академии наук Института молекулярной биологии им. РАН
Официальные оппоненты:
Доктор биологических наук, профессор,
академик Юрий Викторович Ильин
Доктор биологических наук, профессор Юрий Федорович Богданов
Доктор биологических наук, профессор Владимир Юрьевич Поляков
Ведущая организация Учреждение Российской академии наук
Институт цитологии и генетики СО РАН
Защита состоится " " 2010 г. в " " часов
на заседании Диссертационного совета Д 002.235.01 при Институте
молекулярной биологии им. РАН по адресу:
С диссертацией в виде научного доклада можно ознакомиться в библиотеке Института молекулярной биологии им. РАН.
Диссертация в виде научного доклада разослана " " 2010 г.
Ученый секретарь
Диссертационного совета
кандидат химических наук
общая характеристика работы.
Актуальность проблемы. Цитогенетика растений менее чем за столетие прошла путь от описания числа и морфологии хромосом в геноме и кариотипе вида (в понимании Г. Винклера, и ) до молекулярного кариотипирования (идентификации хромосом с помощью физического картирования на них различных ДНК-зондов методами флуоресцентной гибридизации in situ). Важнейшим этапом в развитии цитогенетики растений стало внедрение в 70-е годы прошлого столетия в широкую практику Т. Касперсоном, K. Воза и другими учеными методов диффренциального окрашивания хромосом (бэндинга). Метод С-бэндинга, основанный на выявлении участков конститутивного гетерохроматина, на долгие годы занял ведущее место в исследованиях хромосом растений, что связано с чрезвычайной обогащенностью их геномов повторяющимися последовательностями ДНК. В 70-80-х г. г. прошлого века были изучены рисунки С-бэндинга хромосом у многих видов возделываемых и дикорастущих растений. Следующий этап в развитии цитогенетики растений, приведший к серьезному расширению ее возможностей, связан с разработкой и началом широкого использования в 80-90-х годах прошлого столетия флуоресцентной гибридизации in situ (FISH). В качестве маркерных ДНК-зондов начали применять участки генов рибосомных РНК и другие ДНК-последовательности. В результате за прошедшие десятилетия был накоплен огромный багаж знаний, касающихся особенностей хромосомной организации геномов растений, получены важнейшие сведения об их изменчивости, происхождении и эволюции, что послужило базисом для разработки цитогенетических основ селекции. Стал возможен многопараметрический сравнительный анализ кариотипов в природных популяциях, а также у сортов и линий культурных растений и их гибридов. Цитогенетический мониторинг селекционных образцов, исследование селекционной истории и контроль чистоты сортов прочно вошли в практику селекции. В это же время возник широкий спектр методов молекулярного анализа генома, который также внедрился в генетические исследования многих культурных видов растений и их дикорастущих сородичей, а также в селекционную практику. Объединение цитогенетического и молекулярного подходов открыло новые перспективы для изучения геномов растений. В частности, физическое картирование последовательностей ДНК на хромосомах является необходимым этапом при тотальном секвенировании крупных эукариотических геномов.
Однако большинство из перечисленных выше исследований были выполнены на объектах, метафазные хромосомы которых имеют размеры не менее 5 мкм (большинство злаков, лилейные, многие бобовые). Это связано с тем, что изучение рисунка дифференциального окрашивания небольших хромосом (длиной 1-4 мкм) часто представляет значительные трудности вследствие недостаточной разрешающей способности светооптического микроскопа. Кроме того, геномы мелкохромосомных растений, как правило, имеют небольшие размеры (на один-два порядка меньше, чем геномы злаков или лилейных) и содержат сравнительно мало повторяющихся последовательностей ДНК различных классов, а, следовательно, и конститутивного гетерохроматина. Это обуславливает бедность рисунков С-бэндинга мелких хромосом, что еще в большей степени затрудняет их распознавание. В связи с этим, для изучения кариотипов растений, имеющих небольшие хромосомы, возникла необходимость разработки новых подходов, повышающих разрешение методов хромосомного анализа, а также поиск дополнительных хромосомных маркеров. Важность решения этой задачи связана с тем, что кариотипы значительного числа хозяйственно-ценных культур: технических, масличных, овощных и т. п. представлены хромосомами малых размеров. Для большинства таких растений задача соотнесения молекулярной, генетической и цитологической классификаций хромосом остается весьма актуальной. Особое место среди таких растений занимает лен – культура, имеющая для нашей страны не только большое хозяйственное, но и стратегическое значение. Именно поэтому исследование хромосомной организации генома этого ценного растения и его дикорастущих сородичей представляет собой задачу первостепенной важности.
Цель работы.
Разработка новых и усовершенствование существующих методов молекулярно-цитогенетического исследования для повышения разрешающей способности анализа геномов растений с хромосомами малых размеров или малоинформативным рисунком дифференциального окрашивания. Изучение кариотипов таких растений (низших и высших) с целью идентификации индивидуальных хромосом, выявления структурных перестроек, изучения внутривидового хромосомного полиморфизма, а также сравнения геномов родственных видов для уточнения их таксономического статуса и филогенетических взаимосвязей.
Основные задачи исследования:
1. Разработка комплекса методов, повышающих разрешающую способность анализа хромосомной организации геномов растений с хромосомами малых размеров или малоинформативным рисунком С-бэндинга на модельных объектах (человек, ячмень, пшеница) и объектах исследования (ромашка, лен, горох).
2. Поиск дополнительных молекулярных и цитогенетических маркеров для FISH-картирования хромосом и геномов растений с хромосомами малых размеров или малоинформативным рисунком С-бэндинга:
· применение универсальных маркеров – генов рибосомных РНК и теломерной последовательности ДНК,
· маркеры из тотальной геномной ДНК,
· оптимизация методов выявления флуорохромного бэндинга на хромосомах растений после процедуры FISH.
3. Повышение информативности хромосомного анализа с помощью последовательного или одновременного сочетания несколько методов выявления на хромосомах различных молекулярных и цитогенетических маркеров, а также применения специализированных компьютерных технологий и методов геномного анализа.
4. Исследование с помощью разработанных подходов кариотипов растений с хромосомами малых размеров или с неинформативным рисунком дифференциального окрашивания с целью точной идентификации их хромосом, выявления структурных перестроек, определения родственных взаимоотношений, уточнения таксономического статуса и филогенетических взаимосвязей изученных видов. Особое внимание предполагается уделить растениям, имеющим хозяйственную ценность (хлопчатник, лен, горох, ромашка) или используемым в качестве модельных объектов.
Научная новизна.
Экспериментальная часть работы представляет собой исследование приоритетного характера, так как основные результаты получены впервые и/или на объектах, не исследовавшихся ранее.
На модельном объекте – ячмене разработана модификация метода, позволяющая выявлять рисунок ранней репликации хромосом у растений. Показано, что у растений, также как и у животных, наблюдается высокая консервативность последовательности репликации районов хромосом в родственных геномах. Подобно млекопитающим, у растений выявляется сходство рисунков ранней репликации и G-подобного окрашивания хромосом после обработки трипсином.
Разработана методика получения препаратов митотических метафазных хромосом низкой степени конденсации с помощью предобработки делящихся клеток интеркалятором ДНК 9-аминоакридином. Установлено, что для достоверного анализа хромосомного полиморфизма по С-блокам длина недоконденсированных хромосом не должна более чем в три раза превышать средний размер этих же хромосом, получаемых без использования интеркалятора. Методика существенно расширяет возможности анализа хромосом человека и растений, особенно мелкохромосомных видов, а также видов с неинформативным рисунком С-окраски хромосом.
Обнаружено, что окрашивание недоконденсированных хромосом растений ацетоорсеином позволяет выявлять высокоразрешающий G/R-подобный рисунок дифференциального окрашивания (OR-бэндинг), содержащий значительно большее число сегментов по сравнению с С-окраской. Установлено, что OR-бэндинг хромосомо - и видоспецифичен и хорошо воспроизводим. С помощью последовательного OR-С/DAPI-окрашивания хромосом льна и гороха впервые продемонстрировано, что ацетоорсеином не окрашиваются прицентромерные, крупные интеркалярные и теломерные гетерохроматические районы хромосом.
Показано, что локализация с помощью FISH на хромосомах генов рибосомных РНК позволяет использовать их в качестве эффективных дополнительных молекулярно-цитогенетических маркеров для идентификации мелких хромосом и хромосомных перестроек в геномах растений.
Продемонстрирована перспективность использования для растений подходов, разработанных с целью исследования генома человека. Показана принципиальная возможность получения NotI-библиотек из небольших геномов мелкохромосомных растений и их использования для изучения этих геномов и поиска молекулярных маркеров хромосом.
У мелкохромосомных растений использование собственной геномной ДНК в качестве зонда для FISH позволяет выявлять на хромосомах рисунки гибридизации, сходные с C-бэндингом.
Установлено, что применение двух новых АТ-специфичных флуоресцентных соединений – димерных бис-бензимидазолов – позволяет выявлять более контрастные рисунки дифференциального окрашивания по сравнению со стандартно используемыми флуорохромами DAPI и Hoechst 33258. Это не только расширяет возможности точной идентификации хромосом и хромосомных перестроек, но и FISH-картирования на них различных зондов.
По рисункам ранней репликации впервые идентифицированы хромосомы в диплоидном A1 геноме дикорастущего хлопчатника Gossipium herbaceum var. africanum и аллотетраплоидном (AD)2, геноме тонковолокнистого хлопчатника G. barbadense, а также проведено разделение хромосом (AD)2 генома на Ab и Db субгеномы. Обнаружено сходство рисунков ранней репликации хромосом в родственных геномах.
С применением 9-АМА впервые проведено морфометрическое исследование хромосом одноклеточных красных водорослей и показано, что число хромосом может быть использовано для уточнения таксономии класса Cyanidiophyceae (Rhodophyta).
По рисунку С-бэндинга повышенного разрешения (после 9-АМА предобработки) проведена идентификация хромосом в кариотипах ромашки аптечной и двух родственных видов ромашек. Установлено, что Matricaria inodora является автотетраплоидом, субгеномы которого сходны с геномом M. maritima, а M. chamomilla имеет родственный геном.
По рисункам С/DAPI-бэндинга идентифицированы все хромосомы гороха, включая морфологически неразличимые при равномерном окрашивании 1 и 2 пары гомологов. Это позволило провести исследование внутривидового полиморфизма гетерохроматических районов хромосом, а также обнаружить двойную транслокацию с одновременной перицентрической инверсией у линии гороха с перестроенным кариотипом.
Сравнительное исследование геномов двухромосомных злаков из родов Zingeria и Colpodium по молекулярно-цитогенетическим маркерам хромосом и последовательностям внутренних транскрибируемых спейсеров генов 45S рРНК (ITS1 и ITS2) позволило установить, что эти два близких рода имеют монофилетическое происхождение.
Впервые с помощью комплекса высокоразрешающих модификаций методов С/DAPI-бэндинга и одновременного FISH-картирования на хромосомах генов 26S и 5S рРНК изучены геномы возделываемого вида льна (Linum usitatissimum L.) и 25 дикорастущих видов из 6 секций рода Linum L., произрастающих в Евразии и Африке. Установлены хромосомные числа у L. squamulosum Rudolphi (2n = 18), L. komarovii Juz. (2n = 18), L. thracicum Degen. и L. czernjajevii Klokov; уточнены хромосомные числа у L. decumbens Desf ( 2n = 16), L. narbonense L. (2n = 28), L. stelleroides Planch (2n = 18), L. pallescens Bunge (2n = 18). Впервые в кариотипах видов секции Syllinum обнаружены В-хромосомы и изучена их структура и установлено, что число А-хромосом у этих видов равно 28.
По сходству кариотипов, все изученные виды льнов разделились на 8 групп, что совпало с распределением этих видов на кластеры по данным RAPD-анализа. Результаты молекулярно-цитогенетического анализа подтвердили обособленность секции Adenolinum от секции Linum, выявили геномное сходство видов секций Adenolinum и Stellerolinum, а также показали обособленное положение видов L. narbonense L. и L. nodiflorum L. от других видов льна. Проведенный анализ позволил уточнить таксономический статус некоторых видов секций Linopsis, Syllinum, Linum и Adenolinum. Проведена реконструкция возможных филогенетических взаимосвязей культурного и дикорастущих видов льна.
Теоретическая и практическая ценность работы.
Впервые на основании использования цитологических маркеров проведена полная идентификация хромосом у двух видов хлопчатников. Предложенная нами классификация хромосом хлопчатников, остается единственной, и по сей день используется всеми авторами, изучающими геномы возделываемых видов хлопчатников и их дикорастущих сородичей.
Выявлены новые кариологические характеристики видов, входящих в состав родов Galdieria, Cianidium, Gossypium, Matricaria, Zingeria, Calpodium, Linum, позволяющие уточнить их таксономический статус и происхождение.
Полученные результаты могут быть применены для ускорения селекционного процесса, направленного на получение новых высокопродуктивных и устойчивых сортов льна, гороха и ромашки аптечной. Результаты также могут быть использованы в геномных исследованиях льнов секции Syllinum, которые являются продуцентами биологически активных соединений, применяемых в фармакологии в качестве противоопухолевых препаратов.
Применение интеркалятора ДНК 9-АМА и новых флуоресцентных красителей (димерных бис-бензимидазолов) не только расширяет возможности исследования хромосомной организации геномов ценных сельскохозяйственных культур, но и может существенно повысить точность цитогенетического анализа и картирования хромосом человека, что особенно важно для проведения сложной перинатальной цитогенетической диагностики плода.
Сведения, полученные о геномах видов рода Linum, заложили цитогенетические основы для объединения генетической и цитологической классификаций хромосом и тотального секвенирования генома L. usitatissimum L.
Результаты исследования могут быть также использованы в курсах лекций и лабораторных работах по цитогенетике в средних и высших учебных заведениях.
Апробация работы.
Результаты, полученные в данной работе, представлены на следующих научных конференциях: VI International Barley Genetic Symposium, 1991; 17 International Congress of Genetics, 1993; II Российский симпозиум "Новые методы биотехнологии растений", 1993; 26 Annual Meeting of American Society of Cell Biology, 1994; V (1996),VIII (1998) и XI (1999) Конференции "Геном человека"; V International Oat Conference & VII International Barley Symposium, 1997; XI(1997), XII(1999), XV(2005) Всероссийские совещания "Структура и функции клеточного ядра"; XIX(1998), XX(2000), XXIV (2008) International Congress ISAC, II (1997), III (1999), IV (2001) VII(2007) и VIII(2009) Международные симпозиумы "Новые и нетрадиционные растения и перспективы их практического использования"; XIII International Genome Sequencing and Analysis Conference, 2001; Международный симпозиум "Молекулярные механизмы генетических процессов и биотехнология", 2001; International Polyploidy Conference, 2003; III(2004) и V(2009) Съезды ВОГиС; IV International Conference BGRS, 2004; Международная конференция "Молекулярная и прикладная генетика", 2005; Международная научная конференция "Современные проблемы генетики", 2005; V Международное совещание по кариологии, кариосистематике и молекулярной систематике растений, 2005; Human Genome Meeting, 2001 и 2006; 11th World Congress on Advances in Oncology and 9th International Symposium on Molecular Medicine, 2006; Международная конференция "Генетика в России и мире", 2006; 16th International Chromosome Conference (ICC), 2007; Международная конференция "Генетические ресурсы культурных растений в XXI веке: состояние, проблемы, перспективы", 2007; II Научно-практическая конференция "Современные тенденции в селекции и семеноводстве овощных культур. Традиции и перспективы", 2008; II съезд Общества клеточной биологии совместно с юбилейной конференцией, посвященной 50-летию ЦИН РАН, 2007; Симпозиум памяти "Хромосомы и эволюция", С-Петербург 2008; VI(2008), V(2009) Международной научной конференции "Факторы экспериментальной эволюции организмов"; International Conference "Chromosome, 2009".
Публикации. По материалам диссертации опубликовано 52 статьи (среди которых три обзора) в международных и отечественных рецензируемых научных журналах, раздел в книге, 19 статей в сборниках и около 60 тезисов.
Материалы и методы.
Семена ячменя для исследования получены из коллекции Лаборатории генетики растений ИОГен РАН, ромашек – из коллекции ВИЛАР РАСХН, гороха – из коллекции кафедры генетики МГУ и из коллекции ВНИИСХМ, цингерии, кальподиума и катабразеллы – из Лаборатории кариосистематики БИН РАН, хлопчатников – из коллекции Иолотанской опытной станции ВИР РАСХН, льнов – из коллекций ВНИИЛ РАСХН, ИГиЦ НАН Беларусии, Института ботаники НАН Украины и Генного банка Института генетики растений и исследования возделываемых растений г. Гетерслебена (Institute of Plant Genetics and Crop Plant Research, Gatersleben, Germany).
Для исследования хромосом растений были разработаны методы выявления рисунков ранней репликации хромосом и искусственной задержки конденсации митотических хромосом, а также модифицированы методики OR-, С-, DAPI-, CMA - и Ag-ЯОР-дифференциального окрашивания хромосом. G-подобное окрашивание хромосом после обработки трипсином проводилось согласно уже известной методике [Friebe et al., 1996]. Геномная ДНК выделялась стандартным СТАБ-методом [Murray, Tompson, 1980]. Клонирование фрагментов 26S и 5S рДНК, выделенных с помощью ПЦР из генома Linum austriacum L., проводилось в клетках Escherichia coli (XL 10-Gold Ultracompetent Cells, Stratagene) с использованием вектора pTZ57R (Fermentas, St. Leon-Rot, Germany) согласно протоколу производителя. Для получения геномных NotI библиотек и их анализа, определения количества ДНК в расчете на 1С, проведения высокоразрешающего RAPD-PCR-анализа в полиакриламидном геле, анализа последовательностей внутренних транскрибируемых спейсеров ITS1 и ITS2 генов 45S рРНК применены методики, разработанные ранее [Бойко и др., 1985; Kashuba et al., 1999; Родионов и др., 2005; Зеленина и др., 2006]. Для поиска гомологий сиквенсов клонированных последовательностей с имеющимися в базах данных использованы NCBI Blast и Arabidopsis thaliana WU-BLAST2. Для гибридизации на хромосомах геномной ДНК, клонированных геномных фрагментов, генов рибосомных РНК и синтезированной меченной флуорохромом олигонуклеотидной теломерной последовательности арабидопсиса (CCCTAAA)3–С6-Cy3 (БиоЧип, ИМБ РАН) специально были разработаны различные модификации метода FISH. Исследование проводили с помощью систем цитогенетического анализа (микроскоп, ССD-камера), а также специализированных программ хромосомного анализа ImageJ 1.42 (National Institutes of Health, USA), ВидеоТестКарио 1.5 и 3.0 (ВидеоТест, Сантк-Петербург, РФ).
Основное содержание работы.
Глава 1. Подходы к идентификации мелких хромосом в геномах растений.
Знание любого генома неполно без исследования его хромосомной организации. У растений такая работа обычно начинается с изучения числа и морфологии метафазных хромосом в митозе с последующей идентификацией гомологичных пар. Существует несколько возможностей решения задач идентификации и исследования хромосом в кариотипах растений. Как правило, для каждого конкретного кариотипа необходимо подбирать наиболее эффективные методы анализа и последовательность их применения. Кроме того, нужны способы, позволяющие быстро и с наименьшими потерями информации провести регистрацию изображений хромосом и их объективное исследование. Это легко разрешимо в век компьютерных технологий, которые позволяют быстро получить цифровое изображение хромосом, выявить максимальное число хромосомных маркеров и провести анализ хромосом в интерактивном режиме, т. е. создать объектспецифичный алгоритм изучения хромосом.
При исследовании мелких хромосом со сходной морфологией гарантированный результат можно получить только при комплексном применении всех вышеперечисленных подходов. Именно такой эффективный подход к изучению кариотипов растений с небольшими хромосомами представлен ниже. Подход включает комплекс высокоразрешающих молекулярно-цитогенетических методов исследования, позволяющих получать достаточное число маркеров для распознавания и сравнительного изучения мелких хромосом или хромосом с малоинформативным рисунком С-бэндинга.
1.1. Рисунок ранней репликации хромосом растений.
Поиск новых методов, позволяющих получать рисунок дифференциального окрашивания высокого разрешения, был начат нами с методики выявления репликативного бэндинга, основанной на асинхронности репликации ДНК хромосом в течение S-фазы. В основе этого метода лежит включение в процессе репликации ДНК аналогов оснований с последующей их детекцией. В результате, на хромосомах млекопитающих выявляется высокоразрешающий так назваемый "репликативный" бэндинг. У млекопитающих процесс синтеза ДНК имеет двухфазный характер, поэтому в хромосомах можно выявлять рисунки, соответственно, ранней и поздней репликации. У человека рисунок ранней репликации сходен с G-, а поздней – с R - и С-бэндингом при использовании так называемого FPG (Fluorochrom Plus Giemsa) метода выявления репликативного бэндинга [Захаров и др., 1982]. Применительно к хромосомам растений мы адаптировали вариант метода выявления рисунков ранней репликации хромосом - BrdU-Hoechst-Giemsa-бэндинг. Он основан на встраивании бромдезоксиуридина (BrdU) в ДНК в начале S-периода ее синтеза и последующим разрушением участков ДНК со встроенным аналогом с помощью облучения ультрафиолетовым светом [Захаров и др., 1982]. При этом участки хромосом, реплицирующиеся в середине и конце S-фазы, выделяются как темно окрашенные (Giemsa-положительные) сегменты (бэнды). В этом варианте методики используется обработка BrdU в высокой концентрации, что блокирует большинство делящихся клеток корневой меристемы в начале S-фазы. После снятия BrdU-блока, в первом митотическом делении на хромосомах выявляется рисунок ранней репликации.
Для отработки методики в качестве модельного объекта было выбрано крупнохромосомное растение – ячмень (Hordeum vulgare L.), хромосомы которого имеют длину 6-12 мкм. У этого модельного объекта нам удалось получить рисунок ранней репликации, который был специфичен для каждой из 7 гомологичных групп хромосом (Рис. 1).

Рис. 1. Кариограмма хромосом H. vulgare. В каждой гомологичной группе (номера 1-7) хромосомы расположены по типам окрашивания слева направо:
– рисунок поздней репликации хромосом (по данным Uozu et al., 1997);
– рисунок ранней репликации хромосом;
– рисунок G-подобной окраски (окрашивание раствором Giemsa после обработки трипсином);
– рисунок С-окраски хромосом.
Ранее считалось, что у растений асинхронность репликация ДНК в течение S-периода выражена в меньшей степени, чем у млекопитающих [Cortes et al., 1980; Kakeda, Yamagata, 1991]. Однако, как показали наши исследования, асинхронность репликации ДНК в течение S-периода у растений может проявляться достаточно отчетливо. При этом на хромосомах ячменя, как и на хромосомах человека [Захаров и др., 1982], рисунок поздней репликации хромосом в целом совпадает с рисунком С-окраски [Uozu et al., 1997], а рисунок ранней репликации хромосом – с рисунком G-подобного дифференциального окрашивания, выявляемого на хромосомах после обработки трипсином (Рис. 1). Этот факт еще раз подтверждает единство основных принципов хромосомной организации геномов высших эукариот.
Большое число сегментов, которые выявил на хромосомах ячменя метод репликативного бэндинга, побудило нас попытаться использовать этот подход для изучения мелких хромосом растений с целью идентификации этих хромосом по рисункам ранней репликации (См. раздел 2.1).
1.2. Исследование недоконденсированных хромосом.
Другой подход, который используют при изучении кариотипов растений с малыми размерами метафазных хромосом – это их анализ в прометафазе или профазе, когда отчетливо виден рисунок дифференциальной конденсации хромосом, который можно применять для их идентификации. Однако, этот метод не получил широкого распространения, поскольку получение большого количества прометафазных и профазных хромосом связано с рядом методических сложностей [Fukui, Mukai, 1988].
Более широкие перспективы имеет подход, основанный на искусственном торможении процесса конденсации хромосом за счет встраивания молекул интеркаляторов между парами оснований молекулы ДНК. К числу ДНК-интеркаляторов, задерживающих конденсацию хромосом, относятся такие широко применяемые флуоресцентные красители, как этидий (ЭБ) или акридиновый оранжевый (АО) [Зеленин, 1967; Zelenin, 1999]. Введение интеркаляторов в делящиеся клетки приводит к увеличению продолжительности профазы и метафазы и накоплению в этой стадии большого числа клеток с недоконденсированными или искусственно "удлиненными" хромосомами. На таких хромосомах разрешающая способность любого дифференциального окрашивания повышается.
С целью замедления конденсации хромосом растений мы использовали предобработку делящихся клеток ДНК-интеркалятором 9-аминоакридином (9-АМА). При выборе этого агента мы исходили из простоты его структуры (отсутствии множественных боковых групп), а также низкой константы связывания 9-АМА с ДНК. Эти особенности 9-АМА гарантируют его легкое удаление из хромосом при фиксации и последующих обработках, а, следовательно, использование 9-АМА не препятствует применению в дальнейшем различных методик дифференциального окрашивания хромосом.
1.2.1. Метод задержки конденсации митотических хромосом с помощью 9-АМА.
Отработку метода задержки конденсации хромосом с помощью 9-АМА проводили на хромосомах человека, поскольку степень их конденсации легко оценивать по числу G-бэндов на гаплоидный набор, и на хромосомах ромашки аптечной (Matricaria chamomilla L.), степень конденсации которых оценивали по числу OR-бэндов. Для сравнения использовали обработку этидием, которую часто применяют для задержки конденсации хромосом человека и животных и получения рисунка дифференциального окрашивания более высокого разрешения. Было исследовано влияние 9-АМА и этидия в различных концентрациях (от 0,1 до 10,0 мкг/мл) на задержку конденсации хромосом. Установлено, что для получения сходных эффектов задержки конденсации хромосом требуется существенно более низкая концентрация 9-АМА, чем этидия.
Добавление в культуру лимфоцитов крови человека 9-АМА до конечной концентрации 0,5-1,0 мкг/мл или этидия - до l,0-5,0 мкг/мл за час перед фиксацией давало возможность получать значительное число метафаз с длинными хромосомами и с небольшим числом хромосомных наложений на препаратах. В этих условиях средняя длина хромосом увеличивалась на 70-90% по сравнению с хромосомами, не обработанными 9-АМА. При одинаковой степени разрешения рисунков G‑бэндинга их контрастность, после обработки 9-АМА, оказалась заметно выше, чем после обработки этидием. Этот факт позволяет полагать, что использование предобработки 9-АМА весьма перспективно для повышения степени разрешения различных типов дифференциального окрашивания хромосом человека при проведении высокоточного цитогенетического анализа.
Для ромашки аптечной оптимальные условия предобработок интеркаляторами, приводящих к достаточному удлинению хромосом и выявлению четкого OR-бэндинга на всех хромосомах, были определены как 4-10 мкг/мл для этидия и 0,5-2,0 мкг/мл для 9‑АМА за 12 часов до фиксации при температуре +26°С. Уменьшение концентрации и времени обработки интеркаляторами приводило к снижению эффекта задержки конденсации хромосом. Увеличение концентраций 9-АМА и этидия и/или длительности обработки этими реагентами приводило к значительному увеличению длины хромосом и числа хромосомных наложений, а также к снижению числа метафазных пластинок на препаратах.
Таким образом был разработан протокол получения препаратов с большим числом "удлиненных" метафазных хромосом, основанный на использовании 9-АМА. Установлено, что обработка 0.5-1.0 мкг/мл 9-АМА в течение часа до фиксации клеток приводит к существенному возрастанию разрешающей способности G-окрашивания хромосом человека с 400 до 850 G-бэндов на гаплоидный набор. Для ромашки оптимальные условия обработки составляли 0.5-1.0 мкг/мл 9-АМА и в течение 12-24 часов до фиксации корневых меристем. При таких условиях 9-АМА не подавляет митотическую активность клеток меристемы, а длина хромосом достаточна для получения высокоразрешающего OR-бэндинга (250-260 бэндов на гаплоидный набор).
Обнаружено, что 9-АМА в качестве интеркалирующего агента имеет ряд существенных преимуществ по сравнению с этидием:
· достижение более сильного деконденсирующего эффекта при более низких концентрациях агента;
· легко отмывается фиксатором и поэтому не флуоресцирует на хромосомных препаратах и не мешает последующему применению различных методов флуоресцентного дифференцильного окрашивания и FISH;
· на хромосомах выявляется более четкий и контрастный рисунок бэндинга.
С учетом всех достоинств в дальнейшей работе для задержки конденсации митотических хромосом с целью повышения разрешающей способности хромосомного анализа мы применяли только 9-АМА.
1.2.2. Повышение разрешающей способности дифференциального окрашивания хромосом.
Методика задержки конденсации митотических хромосом была апробирована для повышения разрешающей способности OR - и С-бэндинга хромосом разных размеров у высших растений (лен, ромашка, горох, цингерия). Кроме того, использование 9-АМА впервые дало возможность визуализировать и исследовать с помощью светового микроскопа хромосомы одного из самых примитивных низших растений – красных одноклеточных водорослей. Установлено, что для всех исследованных объектов оптимальной является одна и та же концентрация 9-АМА – 0,5-1 мкг/мл. Все изложенное позволяет рекомендовать использование этого интеркалятора для получения высокоразрешающего дифференциального окрашивания как хромосом растений, так и человека и животных.
1.2.2.1. Определение оптимальных для исследования интервалов длин недоконденсированных хромосом, полученных после предобработки 9-АМА.
 Использование 9-АМА впервые позволило идентифицировать мелкие хромосомы льна посевного (Linum usitatissimum L.) по рисункам С-окраски повышенного разрешения. При проведении этой работы, мы столкнулись с тем, что применение 9-АМА значительно увеличивает вариабельность метафаз по степени конденсации хромосом на препаратах, что особенно заметно для мелких хромосом (Рис. 2).
Использование 9-АМА впервые позволило идентифицировать мелкие хромосомы льна посевного (Linum usitatissimum L.) по рисункам С-окраски повышенного разрешения. При проведении этой работы, мы столкнулись с тем, что применение 9-АМА значительно увеличивает вариабельность метафаз по степени конденсации хромосом на препаратах, что особенно заметно для мелких хромосом (Рис. 2).
Рис. 2. С-дифференциальное окрашивание хромосом льна (L. usitatissimum L.): а – метафазная и б – прометафазная пластинки.
 Это побудило нас осуществить исследование достоверности оценки размеров и положения С-бэндов на "удлиненных" хромосомах. Сравнение вариабельности рисунка С-окраски в зависимости от степени конденсации хромосом было проведено на примере хромосомы 4 льна крупноцветкового (Linum grandiflorum Desf.), содержащей относительно небольшое число С-бэндов. Препараты хромосом льна были приготовлены в двух вариантах: по стандартной методике с использованием колхицина и по предложенной нами методике с использованием 9‑АМА (1мкг/мл на 12 часов перед фиксацией).
Это побудило нас осуществить исследование достоверности оценки размеров и положения С-бэндов на "удлиненных" хромосомах. Сравнение вариабельности рисунка С-окраски в зависимости от степени конденсации хромосом было проведено на примере хромосомы 4 льна крупноцветкового (Linum grandiflorum Desf.), содержащей относительно небольшое число С-бэндов. Препараты хромосом льна были приготовлены в двух вариантах: по стандартной методике с использованием колхицина и по предложенной нами методике с использованием 9‑АМА (1мкг/мл на 12 часов перед фиксацией).
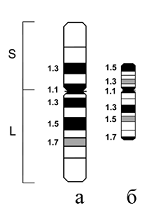
Рис. 3. Идиограмма С- дифференциально окрашенных хромосом: a – ячменя и б – льна.
Для сравнения была взята хромосома 4 ячменя (Hordeum vulgare L.) (Рис. 3). Ячмень был выбран потому, что для этого объекта был предварительно проведен качественный и количественный анализ полиморфизма С-блоков хромосом, и на основе полученных сведений были разработаны основные принципы хромосомной паспортизации по рисункам С-окраски.
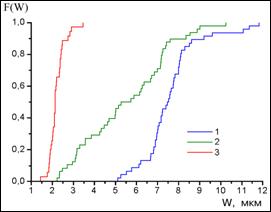 Оказалось, что, несмотря на значительно больший разброс длин хромосомы 4 льна (2,3-7,1 мкм), после обработки 9-АМА по сравнению с необработанными (1,5-2,8 мкм) (Рис. 4), основной рисунок С-окраски мало изменяется. При длине хромосомы от 2,3 до 5,5 мкм обнаружено постоянство размеров и вариабельности крупных гетерохроматических блоков; не меняется также положение на хромосомных плечах небольших интеркалярных С-бэндов. Это позволяет проводить оценку полиморфизма С-блоков в хромосоме 4 льна, подобно тому, как это делается и у ячменя.
Оказалось, что, несмотря на значительно больший разброс длин хромосомы 4 льна (2,3-7,1 мкм), после обработки 9-АМА по сравнению с необработанными (1,5-2,8 мкм) (Рис. 4), основной рисунок С-окраски мало изменяется. При длине хромосомы от 2,3 до 5,5 мкм обнаружено постоянство размеров и вариабельности крупных гетерохроматических блоков; не меняется также положение на хромосомных плечах небольших интеркалярных С-бэндов. Это позволяет проводить оценку полиморфизма С-блоков в хромосоме 4 льна, подобно тому, как это делается и у ячменя.
Рис. 4. Функция распределения длин хромосом ячменя (1), льна после обработки 9-АМА (2), льна без обработки 9-АМА (3).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
