
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Концентрация фитопланктона в слоях повышенной концентрации бывает в 4-55 раз выше фоновой (Ryan et al., 2008). Пространственное распределение зоопланктона отличается столь же выраженной микромасштабной неоднородностью (Haury et al. 1978; Mackas et al.1985; McManus et al., 2005). Те копеподы, которые способны эффективно отыскивать пятна и слои повышенной концентрации и питаться в них, могут обеспечивать себя пищей даже в период ее низкой, в среднем, концентрации. Ctenocalanus citer питается наиболее активно в осенне-зимний период. Его жизненный цикл существенно отличается от циклов рачков, которые прекращают питание при сезонном снижении концентрации фитопланктона.
3.4. Сезонная цикличность питания. Основной пик питания растительноядных копепод в высоких широтах связан с «цветением» фитопланктона. В период низкой концентрации фитопланктона те копеподы, которые остаются активными, дополняют свой рацион за счет других источников пищи, при этом их рацион обычно сильно снижается.
К числу копепод, не питавшихся в зимнее время, из исследованных нами в течение года относятся южный Calanoides acutus (Рис. 5) и северный Calanus hyperboreus (Рис. 6). Остальные антарктические виды, хотя и заметно снижали пищевую активность (кроме Ctenocalanus citer), сохраняли способность потреблять пищу. C. acutus и C. hyperboreus совершают сезонные миграции с большой амплитудой, зимуя на глубине до (Воронина, 1984; Atkinson et al., 1997; Auel, Hagen, 2002; Hirche et al., 2006) в состоянии диапаузы, к которой мы подробнее обратимся позже. Несмотря на уход основной части популяции Metridia gerlachei зимой в глубину (Schnack-Schiel, 1998), активность питания этого вида за счет перехода к хищничеству снижается незначительно. Доля питающихся особей в популяциях Calanus propinquus и Rhincalanus gigas зимой резко падает, но часть животных продолжает питаться. Первый из этих видов, хотя в зимний период и уходит из поверхностного слоя, не погружается столь глубоко, как. C. acutus. Наши исследования показали, что при попадании в более благоприятные условия (биотоп с повышенной концентрацией пищи) C. propinquus начинают активно питаться, в то время как кишечники находившихся в том же «пятне» C. acutus оставались пустыми (Pasternak, Schnack-Schiel, 2001b.). Зимой R. gigas рассредоточен по глубине от 100 до 1000 м (Schnack-Schiel, 1998). Питавшиеся особи не были связаны с пятнами повышенной концентрации. Большая доля неоформленной массы в кишечниках (Рис. 2) предполагает питание микрозоопланктоном и хищничество, что согласуется с мнением ряда авторов (Арашкевич, 1978; Marin, Schnack-Schiel, 1993; Pasternak, 1995). Мелкий рачок Microcalanus pygmaeus, потребляющий фитопланктон и детрит, практически не меняет своего вертикального распределения со сменой сезона, погружаясь лишь на 50-60 м (Schnack-Schiel, 1998). Ядро его популяции расположено под термоклином, и мы полагаем, что этот рачок в основном пользуется оппортунистической тактикой добывания пищи, собирая все доступные ему погружающиеся частицы.
Необычная для растительноядной копеподы тактика обнаружена у Ctenocalanus citer (Рис. 5, Pasternak, Schnack-Schiel, 2007). Его пищевая активность намного выше в осенне-зимний период, чем в весенне-летний. Находясь в верхних слоях воды, рачок питается зимой связанными со льдом водорослями.
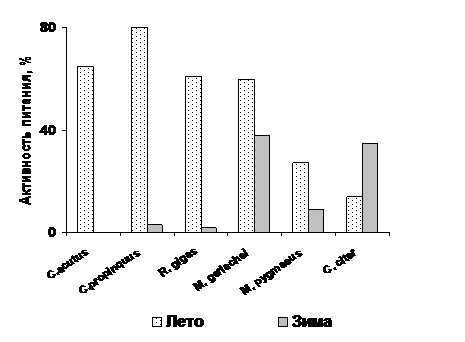
Рис. 5. Сезонные изменения активности питания (% самок и пятых копеподитных стадий с пищей в кишечнике) растительноядных антарктических копепод.
Нам удалось провести почти круглогодичное сравнение активности питания Calanus hyperboreus и C. finmarchicus, совместно обитающих в норвежском фьорде Маланген (Рис. 6). Период питания у C. hyperboreus был намного короче (с марта по июль), чем у C. finmarchicus (с марта по декабрь). Процент питавшихся особей за весь период питания был выше у C. hyperboreus - 80 и 100% у самок и пятых копеподитов против 35 и 52% у C. finmarchicus. Этот факт, как и прямая оценка величины рациона, которая обсуждалась выше, говорит о необычно высокой интенсивности питания C. hyperboreus в период цветения.
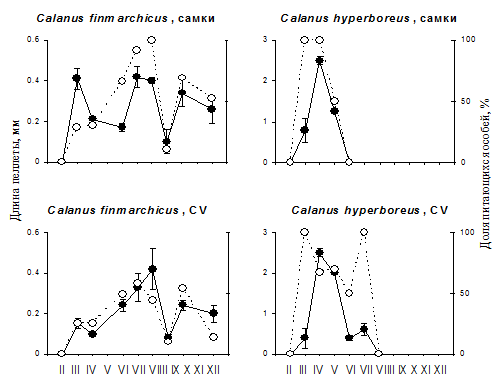
Рис. 6. Сезонные изменеиня активности питания у самок и CV Calanus finmarchicus (левая панель) и тех же стадий C. hyperboreus (правая панедь). Черные кружки и сплошная линия - среднее значение длины пеллеты + SD; белые кружки и прерывистая линия - процент особей с пищей в кишечнике.
х х х
Выделены 4 типа сезонных циклов пищевой активности растительноядных копепод высоких широт с учетом времени начала (по отношению к цветению фитопланктона) и продолжительности периода питания, использованию диапаузы и синхронности при начале и окончании питания (Рис. 7). Тип I (тип «Calanus hyperboreus») характеризуется синхронным и ранним (еще до наступления максимума фитопланктона) началом периода питания. Этот период относительно короток и сменяется ранним переходом к диапаузе. Для этого типа характерна исключительно высокая удельная скорость питания, выше, чем у гораздо более мелких видов растительноядных копепод (Рис. 3). Тип II (тип «C. finmarchicus») – питание наступает в период увеличения концентрации фитопланктона, причем пик питания приходится или на максимум «цветения» или слегка запаздывает. Хотя сезонная активность питания носит пиковый характер, питание с интенсивностью ниже максимальной происходит у части особей в течение почти всего года. Этот тип также предполагает уход в глубину и диапаузу, но позднее, чем у представителей I типа. Тип III (тип «Metridia gerlachei») используется животными, не делающими перерыва в сезонной активности на диапаузу. Активность их питания в осеннее-зимний период снижается незначительно. И, наконец, тип IV (тип «Ctenocalanus citer») – питание происходит в течение всего года. Однако в отличие от большинства растительноядных копепод, максимум пищевой активности которых наблюдается весной и летом, активность питания представителей этого типа достигает максимума в осенне-зимний период. В настоящее время к копеподам, использующим этот тип сезонной цикличности питания, можно отнести лишь C. citer.
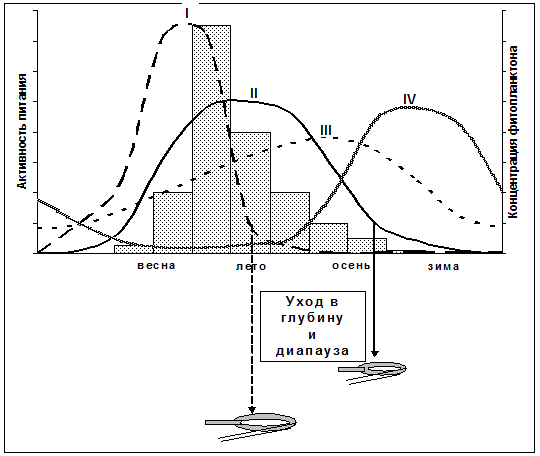
Рис. 7. Сезонные циклы пищевой активности растительноядных копепод высоких широт. I - тип "Calanus hyperboreus" (C. glacialis, Calanoides acutus); II - тип "C. finmarchicus" (C. propinquus, C. glacialis, Rhincalanus gigas); III - тип "Metridia gerlachei" (M. longa, Microcalanus pygmaeus, C. propinquus, Rhincalanus gigas); IV - тип "Ctenocalanus citer"
Глава 4. Репродуктивные стратегии растительноядных копепод высоких широт.
Разнообразие жизненных циклов растительноядных копепод в значительной степени определяется особенностями их размножения. Связь процессов размножения и питания очевидна: самка должна быть обеспечена энергией сверх потребностей базового метаболизма, чтобы в ее организме могли формироваться и созревать ооциты. Начало и продолжительность периода размножения в условиях резко выраженной сезонной цикличности пищевых ресурсов имеют решающее значение, как для продукции яиц, так и для выживания потомства. Совпадение периода размножения рачков с периодом «цветения» фитопланктона и способ обеспечения процесса размножения энергией положены в основу анализа репродуктивных стратегий растительноядных копепод.
4.1. Cезонная цикличность в размножении. Естественно предположить, что у растительноядных копепод будет наблюдаться один сезонный максимум размножения, связанный с периодом цветения фитопланктона. Однако реальная картина гораздо разнообразнее. Используя собственные и опубликованные эмпирические данные, я выделяю основные типы репродуктивной активности копепод.
Изменение доли самок на стадии GS4 и числа зрелых ооцитов в гонадах указывают на два пика размножения у ключевого вида планктона северной Атлантики Calanus finmarchicus. Первый пик связан с размножением перезимовавшей популяции и совпадает с началом сезонного «цветения». Второй, наступающий раньше на шельфе и позже – в открытой части моря, был связан, скорее всего, с размножением новой генерации копепод этого года (Pasternak et al., 2004) и наблюдался на фоне снижения обилия фитопланктона. Однако активность питания C. finmarchicus была в этот период даже выше, чем во время цветения (Рис 5). Возможно, C. finmarchicus в этот период потреблял, кроме фитопланктона, наногетеротрофов и детрит.
Calanus hyperboreus откладывает яйца задолго до «цветения», находясь в глубоких слоях воды (Conover, 1965, 1967; Smith, 1990; Hirche, Niehoff, 1996). Развитие яиц происходит целиком за счет накопленных в предшествующий период питания липидных резервов. По собственным наблюдениям, уже в мае в период цветения C. hyperboreus в Баренцевом море и Маланген-фьорде в северной Норвегии не размножался. Большая часть самок, собранных в поверхностном слое в это время, обладала неразвитыми гонадами (стадия GS 1), меньшая – GS 5 (стареющие самки).
В ранних работах предполагалось, что сезонная периодичность процессов размножения Calanus glacialis сходна с таковой у C. finmarchicus. В дальнейшем было показано, что, по крайней мере, часть популяции C. glacialis откладывает яйца еще до начала цветения, подобно C. hyperboreus. Tourangeau, Runge (1991) связывали начавшийся процесс размножения с питанием подо льдом. Smith (1990) предположила, что источником энергии для раннего размножения служат липидные резервы. Наши наблюдения в проливе Фрама также подтверждают раннее начало размножения у C. glacialis до начала «цветения» при низкой концентрации хлорофилла как в толще воды, так и подо льдом, за счет липидных резервов.
В популяциях антарктических копепод периодичность размножения тоже хорошо выделяется по изменению доли зрелых самок на стадии GS4 в течение года (Schnack-Schiel, 1998; собственные данные). Помимо основных четырех сезонов, автор выделяет переходный период от зимы к весне (или раннюю весну), что объясняется существенными изменениями, происходящими в этот период (с одной стороны, «весна света», а с другой – столь же низкая, как в середине зимы, концентрация хлорофилла). У Calanoides acutus и Calanus propinquus максимальная численность зрелых самок отмечена весной (начало цветения), у Microcalanus pygmaeus и Ctenocalanus citer в летне-осенний период (после цветения), а у Metridia gerlachei летом (в разгар цветения). Самый короткий период размножения из всех этих видов у C. acutus, затем в порядке возрастания следуют C. propinquus, C. citer, M. gerlachei и, наконец, размножающийся практически круглогодично M. pygmaeus. О сезонном изменении доли зрелых самок в популяции Rhincalanus gigas данных нет, но по имеющимся сведениям о растянутом (с весны до поздней осени) периоде размножения (Ommaney, 1936; Voronina, 1970; Воронина и др., 1980; Atkinson, 1991; Marin, Schnack-Schiel, 1993; собственные данные) этот вид можно поместить рядом с M. gerlachei.
Оценки продолжительности и времени начала периода размножения, полученные при определении доли самок со зрелыми гонадами в популяции, говорят о потенциальном репродуктивном сезоне. Реальный период откладки яиц у значительной части копепод ограничен, по-видимому, поступающей в организм самки энергией. Изменения интенсивности продуцирования яиц во времени свойственны всем копеподам, но параметры цикличности различны и связаны с прочими особенностями жизненных циклов (состав пищи и стратегии питания, особенности жиронакопления, диапауза, способы защиты от хищников).
4.2. Связь размножения с питанием. На полевых данных редко удается обнаружить зависимость скорости продукции яиц от концентрации фитопланктона. Отчасти это связано с тем, что копеподы, кроме фитопланктона, используют другие источники пищи. Основную же роль играет запаздывание между формированием яиц и потреблением пищи. Минимальное время, необходимое для перехода энергии пищи в ооциты, у Calanus finmarchicus составляет 2-3 дня (Hirche, 1990; Hirche et al., 1997; Niehoff et al., 1999). У копепод, накапливающих жировые запасы и впадающих в диапаузу, связь между размножением и питанием еще более опосредована. Временной лаг у них может составлять до восьми месяцев. Одновременное использование при размножении энергии, поступающей с пищей, и запасенной в липидах, еще более усложняет картину.
4.2.1. Развитие гонад и созревание ооцитов. Гипотезу о том, что у большинства растительноядных копепод формирование гонад и начальные стадии оогенеза происходит в основном за счет запасенных жиров (Niehoff, Hirche 1996), мы проверили, исследуя созревание гонад у самок Calanus finmarchicus в Норвежском море. Была получена достоверная отрицательная корреляция между степенью зрелости гонад и объемом жира (коэффициент корреляции Спирмэна R= - 0.56, p<0.001). Это показывает, что формирование гонад в отсутствии питания происходит за счет накопленных ранее липидных запасов.
Несмотря на достижение полной зрелости гонад за счет липидов, яйца у Calanus finmarchicus до конца не созревают и вымет не происходит, пока самка не начинает питаться. Питание на завершающей стадии играет роль скорее триггера, чем источника энергии. Так, наши данные указывают, что полностью созревшие ооциты появляются в гонадах C. finmarchicus у побережья северной Норвегии уже в марте, в самом начале увеличения концентрации фитопланктона и крайне низкой активности питания.
Иная картина наблюдается у C. hyperboreus, у которого все стадии формирования яиц осуществляются за счет липидных резервов (Heinrich, 1962; Conover, 1967; Smith, 1990; Hirche, Niehoff, 1996). У C. glacialis первые кладки полностью обеспечиваются внутренними энергетическими запасами, а последующие – за счет питания во время «цветения» фитопланктона. Исследователи расходятся во взглядах на роль раннего размножения (до начала питания) у C. glacialis: Smith (1990) считает, что оно играет определяющую роль в выживании популяции, а Hirche, Kattner (1993) и Melle, Skjoldal (1998) - что эта роль незначительна. Мы вернемся к адаптивному значению раннего размножения в главе 7.
Связь размножения и питания у антарктических копепод гораздо менее изучена. По-видимому, большинство из них нуждается в притоке свежей энергии для завершения оогенеза, однако, возможно, Calanoides acutus, подобно C. glacialis, может начинать размножение, не приступив к питанию, впоследствии увеличивая скорость продукции яиц при увеличении концентрации фитопланктона (Varpe et al., 2007).
4.2.2. Продукция яиц и обеспеченность пищей. Изменчивость связи между продукцией яиц и скоростью питания в зависимости от временных масштабов исследований проанализирована нами в масштабах месяцев, суток, часов на примере Сalanus glacialis и C. finmarchicus из Баренцева моря (Рис. 8, по Pasternak et al., 2002). Скорость питания оценивалась по числу выделенных (в лаборатории) или собранных седиментационными ловушками (в море) фекальных пеллет, продукция яиц - по числу выметанных (в лаборатории) или собранных батометром (в море) яиц.
Возрастание продукции яиц с увеличением потребления пищи при 24-часовых наблюдениях обнаружена и у C. finmarchicus, и С. glacialis, но лишь у первого вида эта зависимость существенна и достоверна. Вероятно, причина различия кроется в том, что С. glacialis в большей степени, чем C. finmarchicus, зависит от энергии запасенных жиров и в
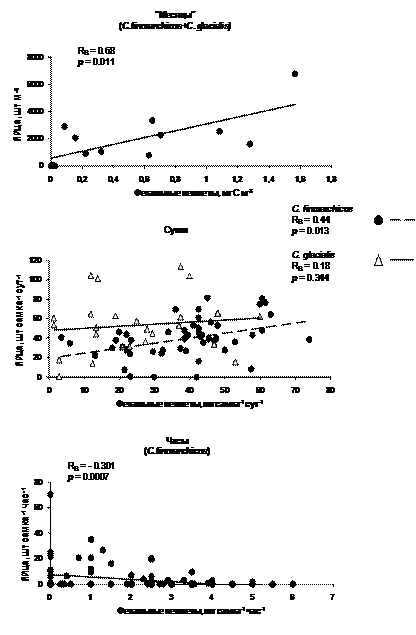
Рис. 8. Зависимость между продукцией яиц и активностью питания копепод в масштабах месяцев, суток и часов. Вверху: одновременно полученные оценки концентрации яиц и пеллет 2-х видов калянусов в марте-августе в Баренцевом море. На двух других графиках: результаты экспериментальных оценок продукции яиц и пеллет.
меньшей - от условий питания непосредственно перед кладкой. При почасовом наблюдении обнаруженная положительная зависимость меняется на отрицательную, т. е, во время откладки яиц самка C. finmarchicus прекращает питание. При осреднении полевых данных по месяцам выявлена четкая положительная связь между двумя процессами (Табл. 1).
Таблица 1. Связь между концентрацией яиц, науплиев и фекальных пеллет Calanus finmarchicus и C. glacialis и концентрацией пищи в слое 0-100 (март, май, июль 1999) в Баренцевом море. Приведены коэффициенты корреляции Спирмэна.
Фекальные пеллеты | Яйца | Науплии | ||||
RS | p | RS | p | RS | p | |
Общий фитопланктон | 0.16 | 0.590 | 0.36 | 0.223 | 0.32 | 0.288 |
«Съедобный» фитопланктон* | 0.63 | 0.020 | 0.59 | 0.032 | 0.66 | 0.014 |
Фитопланктон и простейшие | 0.66 | 0.001 | 0.62 | 0.025 | 0.69 | 0.009 |
Фекальные пеллеты | 0.68 | 0.011 | 0.83 | 0.000 | ||
Яйца | 0.91 | 0.000 |
*В категорию «съедобного» фитопланктона включены клетки размером 5-80 микрон; в нее не попали весьма многочисленные мелкие жгутиковые и Phaeocystis pouchettii
Оценка величины кладки яиц у трех доминирующих видов северного полушария, Calanus hyperboreus, C. glacialis и C. finmarchicus, проведенная до начала сезонного развития фитопланктона и во время «цветения», показала, что в условиях сезонно низкой концентрации хлорофилла лишь C. hyperboreus отличался значительным размером кладки (Рис. 9). Количество яиц в кладке C. glacialis было невелико, а C. finmarchicus вообще не откладывал яиц. В период «цветения», напротив, C. hyperboreus не размножался, а C. finmarchicus приступил к размножению. Величина кладки у C. glacialis заметно возросла.
Периодом, предшествующим «цветению», я называю период с крайне низкой концентрацией хлорофилла (< 0.1 мкг л-1).
Различные типы репродуктивного ответа на возрастание концентрации фитопланктона были выявлены нами на примере двух массовых антарктических видов, Calanoides acutus и Metridia gerlachei на постоянной планктонной станции в декабре 2003 г. в море Уэдделла. По мере развития «цветения» продукция яиц самками C. acutus увеличилась от 14 до 30 экз сут-1 при возрастании доли питающихся самок от 6 до 60% и увеличении числа фекальных пеллет от 33 до 65 экз сут-1. Первые кладки были сделаны в основном за счет липидных запасов, в дальнейшем возрастала роль энергии, вновь поступающей с пищей. Продукция яиц самками M. gerlachei (22-41 экз инд-1 сут-1) варьировала незакономерно на фоне практически постоянной пищевой активности (20-27 пеллет инд-1 сут-1). M. gerlachei питается круглый год, и заметного возрастания активности питания этого вида по мере развития цветения не наблюдалось.
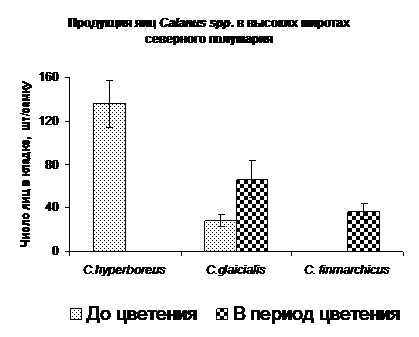
Рис. 9. Продукция яиц копеподами с разными репродуктивными стратегиями в период весеннего "цветения" фитопланктона и в предшествующий "цветению" период.
Зависимость генеративных процессов от обеспеченности пищей проявляется при формировании гонад, созревании ооцитов и продуцировании яиц. У одних копепод весь генеративный цикл может проходить за счет липидных резервов (Calanus hyperboreus), у других поступление новой пищи необходимо лишь на завершающих стадиях формирования яиц (C. finmarchicus), у третьих практически весь цикл должен быть обеспечен поступлением свежей пищи (Metridia gerlachei).
4.3. Типы репродуктивных стратегий. Анализ полученных данных по приуроченности периода размножения копепод к «цветению» фитопланктона, продолжительности репродуктивной активности и связи ее с процессом питания позволил выделить у растительноядных копепод высоких широт три основных типа репродуктивных стратегий.
Не у всех растительноядных копепод размножение приурочено к «цветению» фитопланктона. К числу копепод, не нуждающихся в свежей пищи для начала вымета яиц, относится Calanus hyperboreus, размножающийся задолго до начала «цветения». Этот рачок, зимующий в глубине, откладывает яйца до начала сезонного подъема в поверхностные слои. C. glacialis и Calanoides acutus также могут начинать продукцию яиц до начала питания, однако у этих видов размер кладки при интенсивном питании существенно возрастает. То есть, по признаку начала продукции яиц за счет липидов эти виды сходны с C. hyperboreus, но по признаку ускорения продуцирования с увеличением потребления пищи они ближе к C. finmarchicus. C. finmarchicus не приступает к размножению, не начав питания. Период размножения у копепод этого типа более растянутый, чем у C. hyperboreus, что связано с более продолжительным периодом пищевой активности в поверхностных слоях.
Еще более растянут период размножения у многих более мелких копепод. Так, Microcalanus pygmaeus размножается практически круглый год. Среди возможных причин следует отметить гораздо меньшие, чем у крупных копепод, липидные резервы, что не позволяет им произвести большое число потомков за короткий период. Пищевая пластичность позволяет им находить корм в течение всего года.
х х х
Предлагаемая схема репродуктивных стратегий копепод высоких широт основана на оригинальных и литературных данных (Рис 10). Определяющими параметрами мы считали: а) время начала размножения по отношению к «цветению» фитопланктона, б) продолжительность периода размножения и в) источники энергии для размножения.
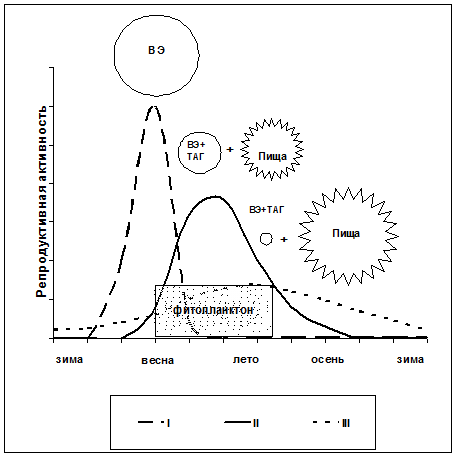 |
Рис. 10. Репродуктивные стратегии растительноядных копепод высоких широт. Над кривыми репродуктивной активности приведены источники энергии для размножения. Круги - запас липидов, многолучевые звезды - энергия пищи. ВЭ - восковые эфиры, ТАГ – триацилглицерины (о составе липидов подробнее – в гл. 6). Примеры: I - Calanus hyperboreus размножается до начала цветения целиком за счет запасенных липидов; II – C. finmarchicus - размножается после начала цветения, за счет как липидов, так и вновь потребленной пищи; III - Microcalanus pygmaeus - размножается круглый год, в основном за счет вновь потребленной пищи.
Репродуктивная стратегия I - размножение с резким сезонным пиком, до цветения фитопланктона. Типичный пример - Calanus hyperboreus. Этой стратегии может следовать C. glacialis и, возможно, Calanoides acutus; она характерна также для представителей р. Neocalanus из Тихого океана (Гейнрих, 2002; Miller, Clemons, 1988). II - размножение с выраженным пиком в период цветения фитопланктона. Ей следуют C. finmarchicus, C. propinquus, Rhincalanus gigas, которые размножаются после начала питания, за счет, как липидов, так и вновь потребленной пищи. III – растянутое размножение, слабо связанное с «цветением» фитопланктона. Этой стратегии следуют копеподы, питающиеся и размножающиеся в течение практически всего года (Microcalanus pygmaeus, Metridia gerlachei, M. longa, Ctenocalanus citer).
Глава 5. Защита от хищников у пелагических копепод: упреждающие и ответные тактики.
Традиционно при исследовании структуры и функционирования планктонных сообществ основное внимание уделялось обеспеченности организмов пищей («bottom-up» эффекты). В последнее время все большая роль отводится влиянию хищников («top-down» эффект, Verity, Smetacek, 1996). Широко признается, что хищничество не только важнейший экологический фактор, но и фактор отбора, определяющий эволюцию целого ряда адаптаций в популяциях жертв, как в поведении (Gliwicz, 1986) и морфологии (Havel, 1987), так и в стратегиях жизненных циклов (Reznick et al., 1990).
Планктонными копеподами питаются как позвоночные (личинки и мальки всех рыб, взрослые планктоядные рыбы, птицы, киты), так и беспозвоночные (хетогнаты, хищные копеподы, медузы, эвфаузииды и др.) животные. Трудно дать в общем виде оценку риска, исходящего от разных групп хищников, тем более, что роль этих хищников изменяется в зависимости от сезона и района. Чаще всего, наибольшая смертность крупных планктонных копепод связана с выеданием рыбами (Brooks, Dodson, 1965; Zaret, 1980; Brodeur et al., 2000; De Robertis et al., 2000; Kaartvedt, 2000; Ciannelli et al., 2004; Leising et al., 2005).
Выделяют 4 группы способов защиты планктона от хищников: химические, морфологические, поведенческие и системные (связанные с жизненным циклом) (Verity, Smetacek, 1996). Другой подход к классификации способов защиты основан на выделении 1) постоянных защитных образований и реакций, 2) возникающих в присутствии хищников («inducible defenses») и 3) отсутствие каких бы то ни было защитных приспособлений (Cohen, Forward, 2005).
Принимая во внимание оба подхода, мы делим применяемые копеподами способы защиты на две крупные категории, «упреждающие» и «ответные», и дальнейшее разделение на морфологические, поведенческие и системные способы проводим внутри этих категорий. Под «упреждающими» мы понимаем все адаптации, не связанные с прямым воздействие хищника, а возникающие под воздействием каких-либо сигнальных факторов. В среде с циклическим закономерным функционированием таким фактором, заблаговременно сигнализирующих о скором увеличении пресса хищников, могут служить изменения освещенности и температуры. «Ответные» защитные реакции возникают лишь в присутствии хищника в ответ на химические, гидродинамические и зрительные стимулы.
5.1. Упреждающие способы защиты от хищников.
Основной морфологический способ защиты планктонных копепод - прозрачность их тела. Наиболее характерным примером поведенческих способов защиты служат суточные вертикальные миграции. Большинство авторов сходятся во мнении, что избегание зрительных хищников (рыб-планктофагов) является основной причиной суточных миграций растительноядного зоопланктона (Zaret, Suffern, 1976; Ohman, 1990; Lampert, 1993), который питается ночью в поверхностных слоях, а днем опускается в глубину, спасаясь от хищников.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
