
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Пелагические экосистемы высоких широт Мирового океана отличаются высокой продуктивностью и одновременно резко выраженной сезонной цикличностью. Растительноядные планктонные копеподы - одно из основных звеньев пищевых цепей в пелагических сообществах, передающее органическое вещество от первичных продуцентов организмам высших трофических уровней (Marshall, 1973; Виноградов, 1977 и др.). Они также играют важнейшую роль в формировании потока органического вещества из трофогенного слоя в глубину (Urrere, Knauer, 1981; Aksnes, Wassmann, 1993). Функцию передаточного звена эти копеподы выполняют в условиях сильной изменчивости среды и ресурса, поскольку период обилия основного корма растительноядного зоопланктона, фитопланктона, в этих широтах ограничен 2-3 месяцами в год. Эти копеподы служат основным кормом многим промысловым рыбам, как молоди, так и взрослым (Cushing, 1984; Kane, 1984; Astthorsson, Gislason, 1997), и адекватная оценка их обилия и продукции необходима при прогнозировании кормовой базы рыб.
Наши суждения о биологических ресурсах пелагиали высоких широт и перспективах их использования зависят от понимания структуры и функционирования основных звеньев этих экосистем (Виноградов 1977; Флинт, Суханова, 2003; Флинт, 2005). Для того чтобы понять закономерности продуцирования пелагических сообществ и колебания обилия массовых видов, необходимо исследовать их жизненные циклы и их адаптивные модификации, позволяющие популяциям гибко приспосабливаться к изменениям условий существования.
Исследование стратегий жизненных циклов - одна из важнейших задач современной экологии (Begon et al., 1990). Каждый стратегия, строго говоря, уникальна, поэтому, чтобы разобраться в этом многообразии, необходимо выделить основные ее параметры. Адаптация копепод к меняющимся условиям идет по пути приспособления жизненного цикла к периодическому изменению абиотических и биотических факторов. Среди характеристик жизненных циклов, представляющих собой закономерное чередование последовательных стадий развития, можно выделить структурные (пространственное распределение, амплитуда миграций, сезонные изменения численности и биомассы, демографическая структура) и функциональные (особенности питания и размножения, роста и развития, накопление резервных веществ, способы переживания неблагоприятных условий). Систематические исследования жизненных циклов массовых видов растительноядных копепод высоких широт начинались с анализа структурных характеристик (Гейнрих, 1962; Воронина, 1972, 1984; Østvedt, 1955; Andrews, 1966; Heinrich, 1962; Voronina, 1970, 1978; Conover, 1988; Marin, 1988a, b; Atkinson 1991; Zmijewska, 1993; Schnack-Schiel, Hagen, 1994). Были выявлены общие черты жизненных циклов растительноядных копепод.
Несмотря на общее сходство, существуют значительные различия в жизненных циклах, как на межвидовом, так и популяционном уровне. Для понимания причин разнообразия жизненных циклов необходимо исследовать эколого-физиологические характеристики жизнедеятельности копепод. Среди них основное внимание привлекают закономерности питания (Арашкевич 1967; Арашкевич, Пастернак, 1984), размножения (Кособокова, 1993; Hirche, 1996; Pasternak et al., 2002; Hirche, Kosobokova, 2003), диапаузы (Алексеев, 1990; Alekseev et al., 2007), жиронакопления (Lee et al., 2006), метаболизма (Ikeda, Mitchell, 1982; Pasternak et al., 1994). Предпринимались попытки объединить структурный и функциональный подходы к исследованию жизненных циклов копепод высоких широт (Conover, Huntley, 1991; Atkinson, 1998; Schnack-Schiel, 2001). Однако новые данные и современные представления о стратегиях жизненных циклов существенно расходятся с предложенными в этих работах классификациями, по-видимому, из-за фрагментарности имевшихся в распоряжении авторов данных и отсутствия единого подхода к анализу жизненных циклов.
Современная функциональная экология все больше переходит от исследований отдельных процессов жизнедеятельности к изучению сложных эколого-физиологических и поведенческих комплексов (Bollens, Frost, 1989; Lima, Dill, 1989; Begon et al., 1990; Михеев, 2006). Для выявления закономерностей формирования жизненных циклов необходимо не только исследовать основные параметры жизнедеятельности, но и проанализировать их взаимодействия. Изучение типов жизненных циклов требует получения сравнимых рядов данных для доминирующих видов, полученных на единой методической основе в течение, по крайней мере, сезонного цикла. К настоящему времени получены новые данные по сезонным изменениям эколого-физиологических характеристик массовых видов копепод высоких широт. В данной работе предлагается обобщение и анализ накопленных данных в рамках представлений о формировании жизненных циклов на основе компромисса между требованиями питания, размножения и защиты от хищников.
Цель работы: Выявить закономерности формирования стратегий жизненных циклов растительноядных копепод высоких широт на основе анализа взаимозависимых процессов питания, размножения и защиты от хищников.
Основные задачи:
- Проанализировать данные по сезонной активности питания и составу пищи массовых видов копепод Арктики и Антарктики. Выявить группы, различающиеся по продолжительности сезона пищевой активности и потребляемой пище.
- На основании оригинальных полевых и экспериментальных данных, а также сведений из литературы, выявить время начала, продолжительность и интенсивность периода размножения у массовых видов высокоширотных копепод. Исследовать зависимость этих характеристик от особенностей питания (продолжительности сезона пищевой активности и потребляемой пищи).
- Проанализировать закономерности оборонительного поведения планктонных копепод, как одного из факторов формирования жизненного цикла. Выявить роль избегания копеподами биотопов с высоким уровнем угрозы в комплексе оборонительного поведения.
- Исследовать основные признаки диапаузы у планктонных копепод: липидный статус и развитие гонад, пищевую и метаболическую активность, глубину обитания. На основе новых экспериментальных и полевых данных уточнить представления о роли диапаузы в формировании жизненных циклов растительноядных копепод.
- Исследовать динамику накопления липидных резервов у копепод в период активного питания и снижения этих резервов при голодании, а также исследовать состав липидов у планктеров, различающихся по характеру потребляемой пищи.
- Выявить основные стратегии жизненных циклов растительноядных копепод, в основе которых лежат разные сочетания ключевых процессов жизнедеятельности: питания, размножения и защиты от хищников.
Научная новизна. Сформулирована концепция формирования стратегий жизненных циклов планктонных копепод на основе компромисса между требованиями питания, размножения и защиты от хищников. Среди многообразия стратегий на основе характера взаимосвязи между основными процессами жизнедеятельности выделено четыре основных типа. Прежде всего, это 2 типа циклов, связанных с избеганием «трофогенной зоны» в неблагоприятный сезон и 2 типа циклов, связанных с приспособлением к ней. Стратегии избегания предполагают, что при сезонном ухудшении условий обитания копеподы уходят на глубину и впадают в диапаузу. Для этих стратегий характерны: короткий (ограниченный весенне-летним сезоном) период размножения и накопление в продуктивный сезон значительных липидных резервов, состоящих преимущественно из восковых эфиров. Стратегии приспособления предполагают, что копеподы не уходят в диапаузу, а остаются активными круглый год. Для этих стратегий характерны: растянутый период размножения и значительно меньшие липидные резервы, которые служат не для переживания продолжительного неблагоприятного периода, а для подстраховки при локальном дефиците корма. У копепод, использующих стратегию приспособления, в качестве липидных резервов в основном откладываются триацилглицерины.
Впервые на единой методической основе определена продолжительность сезона питания у доминирующих видов Антарктики и Арктики. Выявлены характерные сезонные изменения состава их пищи, различающиеся у разных онтогенетических стадий и разных групп видов.
Разработана новая модификация экспресс-метода определения интенсивности питания растительноядных копепод по флуоресценции пигментов в кишечнике с помощью лазерного флуориметра. Предложенный способ позволяет быстро проводить оценку растительных пигментов в кишечнике копепод при малой численности особей (вплоть до одной) в пробе.
Впервые одновременно сделана оценка сравнительной интенсивности дыхания и скорости экскреции в популяциях антарктических копепод, часть которых находилась в активном состоянии в поверхностных слоях, а другая часть находилась в диапаузе на глубине. Если у диапаузирующих особей скорость метаболизма (с поправкой на разницу температуры) в глубине была в 3-10 раз ниже, чем у находившихся в активном состоянии в поверхностных слоях воды особей той же популяции, то у не-диапаузирующих видов такой разницы не наблюдалось.
Впервые прослежены изменения, происходящие в генеративной системе пятых копеподитных стадий копепод в течение всей диапаузы. Показано, что увеличение размеров гонад и их дифференциация происходят на протяжении всего периода диапаузы исключительно за счет энергии накопленных липидов.
Исследовано возникновение диапаузы в популяции копепод под влиянием высокого пресса хищников. В популяции того же вида, существующей в биотопе с низким прессом хищников, диапаузы не наблюдалось.
Основные положения:
Основными эколого-физиологическими детерминантами жизненных циклов растительноядных копепод высоких широт служат: диапауза, пищевая пластичность, разнообразие репродуктивных стратегий, тактики превентивного избегания хищников.
Жизненные циклы планктонных копепод формируются на основе компромисса между требованиями питания, размножения и защиты от хищников. В рамках этой концепции многообразие циклов сводится к четырем основным типам, различающимся по способу переживания неблагоприятного сезона, пищевой пластичности, продолжительности периода размножения и его приуроченности к «цветению» фитопланктона.
Компромисс между размножением, питанием и обороной определяет время и продолжительность размножения. При размножении и активном питании возрастает риск быть замеченным и съеденным хищником. Более «осторожные» копеподы размножаются до цветения фитопланктона, когда потенциальных хищников также еще мало.
Практическое значение. Разработанные в диссертации представления о формировании жизненных циклов копепод важны для анализа потоков энергии в пелагических экосистемах и их изменчивости в связи с колебаниями среды обитания, вызванными изменениями климата и антропогенным воздействием. Выявленные закономерности могут служить основой для совершенствования методов оценки кормовой базы планктоядных рыб. Анализ эколого-физиологических характеристик копепод позволяет понять механизмы изменчивости и разнообразия жизненных циклов. В диссертации разработана концептуальная схема стратегий жизненных циклов растительноядных копепод высоких широт и проанализирована взаимосвязь между основными процессами жизнедеятельности. Показано, что тип жизненного цикла может существенно влиять на эффективность передачи энергии от первичных продуцентов к высшим трофическим уровням, формирование кормовой базы и кормовых миграций рыб. Положения диссертации об эколого-физиологические детерминантах, формировании жизненных циклов копепод на основе компромисса между питанием, размножением и обороной, схема классификации жизненных циклов могут быть включены в курсы лекций и учебные пособия по водной экологии и биоокеанологии для высших учебных заведений.
Апробация работы. Основные результаты работы были доложены на 10 всероссийских/всесоюзных и 15 международных конференциях, а также на семинарах и коллоквиумах лаборатории экологии и распределения планктона Института океанологии РАН, Института морских и полярных исследований им. А. Вегенера (Бремерхафен, Германия), Института полярной экологии (Киль, Германия), университетов Бремена (Германия), Тромсе (Норвегия), Ювяскюля (Финляндия), Глазго (Великобритания).
Публикации. Результаты работы опубликованы в 61 научных работах, в их числе 31 статья в международных журналах.
Структура диссертации. Диссертация состоит из Введения, 7 глав и Выводов. Работа изложена на страницах машинописного текста, включает рисунков и таблиц. Список литературы содержит 325 наименований на русском и иностранных языках.
СОДЕРЖАНИЕ РАБОТЫ
Глава 1. Подходы к изучению жизненных циклов планктонных копепод
Cтруктурные характеристики, в первую очередь, данные по вертикальному распределению и возрастной структуре, полученные в разные сезоны и в разных районах, послужили основой анализа жизненных циклов и оценки продолжительности развития копепод высоких широт (Воронина, 1972, 1984; Ottestad, 1932; Andrews, 1966; Voronina, 1970; Conover, 1988; Marin, 1988a, b; Atkinson 1991; Schnack-Schiel, Hagen, 1994 и др.). В общих чертах жизненные циклы большинства растительноядных копепод высоких широт сходны. Это – интерзональные виды, населяющие в разные периоды своей жизни столб воды от поверхности до глубины м (Виноградов, 1968; Воронина и др., 1978). Их молодые стадии откармливаются на обильном корме в поверхностном слое весной-летом в короткий период «цветения» фитопланктона, развиваются и откладывают жировые запасы. Многие из них осенью погружаются в глубину и впадают в диапаузу, прекращая питание и развитие. Сложившиеся представления о сходстве жизненных циклов растительноядных копепод представляют собой лишь самую общую схему, и конкретные виды, также как популяции того же вида в различных районах, заметно различаются по характеру адаптаций к сезонной цикличности ресурса.
Понимание механизмов, лежащих в основе формирования жизненных циклов копепод высоких широт, требует привлечения не только структурных, но и функциональных (эколого-физиологических) характеристик. По ним накоплен большой материал: питание (Арашкевич, 1969; Воронина, Суханова, 1976; Пастернак, 1984, 1988; Дриц, Пастернак, 1993; Marshall, Orr, 1955; Tande, Båmstedt, 1985; Pasternak, Schnack-Schiel, 2001), продукция яиц (Conover, 1967; Hirche, 1989; Кособокова, 1993; Hirche, Niehoff, 1996; Niehoff et al., 2002.), составу и объемам накапливаемых липидов (Kattner, Hagen, 1995; Hagen, Auel, 2001; Lee et al., 2006), скорости дыхания и других индикаторов метаболизма (Conover, Corner, 1968; Ikeda, Mitchell, 1982; Pasternak et al, 1994). Особую ценность для анализа жизненных циклов представляют круглогодичные данные по эколого-физиологическим характеристикам. Они крайне немногочисленны (Hagen, Schnack-Schiel, 1996; Pasternak, Schnack-Schiel, 2001; Pasternak et al., 2001).
В нескольких работах, анализирующих жизненные циклы копепод, объединяются структурный и функциональный подходы (Conover, Huntley, 1991; Atkinson, 1998; Schnack-Schiel, 2001). Однако полученные обобщения и классификации довольно противоречивы. Одной из основных причин может быть отсутствие единого подхода и критериев к выделению элементов и типов жизненных циклов.
Каждый жизненный цикл по-своему уникален и для того, чтобы упорядочить это разнообразие, мне представляется правильным использовать ограниченное число признаков и критериев. В качестве минимального набора таких критериев для выделения типов жизненных циклов я предлагаю использовать особенности основных процессов жизнедеятельности: питания, размножения и защиты от хищников. Эти виды активности обеспечивают снабжение энергией, сохранение генофонда и выживание популяции. Баланс, или компромисс, между питанием, размножением и защитой от хищников служит основой формирования жизненных циклов планктонных копепод. Процессы жизнедеятельности взаимозависимы, и особенности одного из них накладывают ограничения на функционирование остальных. Например, свойственный данному виду характер питания связан со способом переживания периода недостатка пищи и параметрами репродуктивной активности. Быстрое накопление энергетических резервов вызвано необходимостью раннего ухода из поверхностного слоя в связи с увеличением пресса хищников.
При исследовании стратегий жизненных циклов растительноядных копепод высоких широт крайне важно получение длинных рядов данных по сезонным изменениям эколого-физиологических характеристик. Острый дефицит таких данных, особенно для зимнего и весеннего сезонов, отмечали еще Conover, Huntley (1991). К настоящему времени особенно плохо исследованы в этом отношении мелкие рачки. Помимо высокой численности и значительной роли в трофодинамике пелагических сообществ, данные по этим копеподам могут расширить базу для анализа разнообразия жизненных циклов. Нам удалось существенно пополнить имеющиеся данные результатами длительных наблюдений по питанию, развитию гонад, динамике липидов, а также по эколого-физиологическим характеристикам мелких копепод.
Для понимания механизмов формирования жизненных циклов копепод наиболее важны оценки, полученные на индивидуальном уровне. Мы уделяли особое внимание получению таких оценок, как в полевых, так и экспериментальных исследованиях.
Глава 2. Объект, материал и методы исследования.
Основной объект данного исследования – экологическая группа преимущественно растительноядных копепод (т. е., копепод, не относящихся к облигатным хищникам и использующим фитопланктон как основной источник корма) высоких широт. В строгом смысле слова практически все копеподы - эврифагами. Помимо фитопланктона, они потребляют простейших, детрит, яйца и мелких зоопланктонных животных. Однако в период «цветения» фитопланктона он составляет более 80-90% рациона этих животных (Huntley 1981). Основные материалы были получены мной в Атлантическом секторе Южного океана, северной Атлантике и прилегающих арктических морях, поэтому формирование жизненных циклов я рассматриваю на примере копепод из этих районов.
Питание. Среди традиционно используемых при изучении питания методов, полевых, экспериментальных и математического моделирования, особую ценность для меня представляли методы, позволяющие оценить процессы, происходящие в естественных условиях.
Состав пищи копепод изучался при микроскопическом исследовании содержимого кишечников. Для более точной оценки пищевого спектра проводили фотографирование пищевых остатков. Чтобы оценить сравнительную роль разных пищевых частиц, мы усовершенствовали метод, измеряя размеры пищевых частиц, рассчитывая их объемы и сопоставляя их с общим объемом пищевого комка (Арашкевич, Пастернак, 1984; Pasternak, Schnack-Schiel, 2001a, b, 2007).
Активность питания оценивалась с помощью двух экспресс-индексов, доли в популяции особей, содержащих пищу в кишечнике (Дриц, 1981; Kehayias, 2003), и средней длине пищевой пеллеты (Ayukai, Nishizawa, 1986; Dagg, Walser, 1986). Мы показали, что эти индексы с высокой достоверностью коррелируют между собой и могут служить надежным показателем пищевой активности в естественных условиях (Pasternak et al., 2004).
Скорость потребления пищи и суточные рационы были нами исследованы как in situ с использованием флуоресцентной методики (Дриц и др., 1990; Дриц, Пастернак, 1993; Pasternak et al., 1994; in press), так и в лабораторных инкубационных экспериментах (Пастернак и др., 1981; Пастернак, 1984).
Объем материала. Проведено более 600 микроскопических исследований содержимого кишечников и около 1000 экспресс-оценок активности питания по проценту наполненных особей и длине фекальных пеллет. Скорость потребления пищи была определена по результатам более 1000 измерений флуоресценции пигментов в кишечнике и скорости прохождения пищи, а также 80 инкубационных экспериментов.
Размножение. Из разнообразных методов исследования репродуктивной активности копепод я выбрала оценку скорости продукции яиц при инкубации в лаборатории и определение доли самок со зрелыми гонадами в популяции. Эти подходы, примененные в течение годового цикла, позволили проследить сезонность в репродуктивной экологии копепод, что особенно важно при сравнительном анализе жизненных циклов. Выделение стадий зрелости гонад проводилось согласно классификации (Hirche, Niehoff, 1996; Niehoff, Hirche, 1996), в которую я внесла небольшие изменения, упростив ее и по-иному определив 1-ю стадию (Pasternak et al., 2001). Готовность популяции к размножению определялась по доле самок со зрелыми гонадами (стадия GS4), а максимальный размер кладки – по числу зрелых ооцитов в гонадах (Runge, 1987; Niehoff, Hirche, 1996; Niehoff, 2003; Pasternak et al., 2004). При определении доли зрелых самок мы использовали метод, разработанный (Romeis, 1968) в модификации (Hirche, Niehoff, 1996). Кроме того, мы проследили рост и развитие гонад у пятой копеподитной стадии 2-х массовых видов, Calanus finmarchicus и C. hyperboreus, в течение года. Преимущество этих способов заключается в возможности анализа большого объема материала без экспериментального воздействия на животных.
Объем материала. Всего проведено 450 инкубационных экспериментов в Гренландском, Баренцевом и в море Уэдделла и 40 опытов по оценке интервалов между последовательными кладками (Баренцево море и море Уэдделла). Оценка зрелости гонад проведена в течение годового цикла на 140 экземплярах самок массовых видов копепод из Норвежского моря и моря Уэдделла и 200 экземплярах пятых копеподитов. Соотношение численности полов исследовано у Calanus finmarchicus и C. hyperboreus в течение года, а у антарктических копепод Calanoides acutus, Metridia gerlachei, Microcalanus pygmaeus, Ctenocalanus citer в весенний и в зимний период. Вскрыты гонады у 180 самок C. finmarchicus и по числу зрелых ооцитов оценен размер потенциальной кладки в течение сезона.
Защита от хищников. В морских условиях влияние пресса хищников на миграционную активность и жизненный цикл копепод трудно исследовать с необходимой детальностью. Влияние хищников на миграции и сезонную динамику мы исследовали на примере пресноводных пелагических копепод сем. Diaptomidae, широко распространенных в озерах умеренной и бореальной зоны. Для исследования миграций, изменений в численности и демографической структуре популяций проводился детальный отбор проб по стандартной схеме, а также соотношение особей в активном и пассивном состоянии.
Для оценки сезонных изменений пресса хищников в море использовались имеющиеся в литературе сведения о распределении, обилии и интенсивности питания основных потребителей копепод в водах Арктики и Антарктики (Kellermann, 1986; Hubold, 1992; Zmijewska, Yen, 1993; Dalpadado et al., 2000; Kaartvedt, 2000; Varpe et al., 2005).
Объем материала. Влияние хищников на миграции и сезонный ход популяций растительноядных копепод мы исследовали на примере рачков сем. Diaptomidae из мезотрофных озер: Глубокое (Московская область) и Валлерзее (Австрия). В жизненном цикле Eudiaptomus graciloides из озера Глубокого, где пресс рыб-планктофагов был значителен, прослежены изменения численности, возрастной и вертикальной структуры популяции (более 50 проб), скорости развития старших копеподитных стадий (20 экспериментов продолжительностью от 1 до 4 недель) с мая по ноябрь. В озере Валлерзее, где за 10 лет существенно изменились пресс хищников (молоди окуня) и сезонная динамика популяции Eudiaptomus gracilis, были исследованы демографическая структура (около 200 проб), содержание пищи и жира (более 50 проб), вертикальное и горизонтальное распределение рачков (250 проб).
Диапауза. Предварительное представление о том, что популяция может находиться в состоянии диапаузы, можно получить на основании анализа структурных характеристик (вертикального распределения и возрастной структуры). Однако нахождение на глубине стадий, характерных для диапаузы, не всегда означает, что популяция находится в диапаузе. Необходимо выявить функциональные признаки диапаузы, прежде всего, оценить жировые запасы. Мы оценивали общий запас липидов у копепод тремя способами: гравиметрическим, геометрическим и фотохимическим. Состав запасенных жиров (оценка соотношения разных липидных классов) исследован с помощью жидкостной хроматографии, а состав жирных кислот – газовой хроматографии.
Кишечник копепод в состоянии диапаузы обычно пуст. Если поместить диапаузирующую копеподу в условия высокой концентрации пищи, животное не возобновляет питание в течение, по крайней мере, нескольких дней. Часто диапаузирующие животные обладают пониженной подвижностью и даже не реагируют на прикосновение. Мы использовали эти реакции в экспериментах, чтобы получить дополнительные показатели состояния копепод.
Уровень метаболизма копепод в состоянии диапаузы снижается. Скорость дыхания и экскреции копепод мы определяли при инкубации выловленных из моря животных в замкнутых сосудах по разнице содержания кислорода, соединений азота и фосфора в воде до и после инкубации. Кроме того, потенциальная скорость дыхания оценивалась при определении активности ЭТС.
Объем материала. Объем жирового мешка измеряли в течение годового цикла у самок, 5-х и 4-х копеподитных стадий Calanus finmarchicus и C. hyperboreus (около 2000 экз.) из фьорда в северной Норвегии. Вес и состав экстрагированного жира оценен у пяти видов доминирующих копепод Антарктики во время развития весеннего цветения (55 определений, 10-30 экз. в пробе). На этом же материале определено соотношение липидных классов у разных копепод и состав жирных кислот. Общее количество липидов оценено также у трех антарктических копепод в осенний период (50 определений). Кроме того, количество липидов определялось в активной и диапаузирующей гемипопуляциях пресноводного Eudiaptomus graciloides с весны по осень (80 определений, 20-80 экз. в пробе).
Скорость дыхания и экскреции определена у трех видов антарктических копепод в начальный период погружения этих популяций в глубокие слои (около 100 измерений) и в начале «цветения» фитопланктона (20 измерений).
Глава 3. Трофические адаптации растительноядных копепод к циклическим колебаниям среды.
Способность копепод-фитофагов переходить от питания фитопланктоном к потреблению другого корма связана с их пищевой пластичностью. Копеподы проявляют при питании избирательность, в соответствии с размерами и формой кормовых частиц и их вкусовыми качествами (Frost, 1977; Poulet and Marsot, 1978; Donaghay and Small, 1979; Huntley, 1981). Выбор частиц по размерам и форме связан со строением ротовых придатков и механизмом питания (Арашкевич, 1969; Nival, Nival, 1973, 1976; Boyd, 1976; Huntley, 1981). При изучении питания растительноядных копепод высоких широт мы сосредоточили основное внимание на исследовании пищевых спектров и сезонной изменчивости пищевой активности.
3.1. Формирование пищевых спектров. Исследуя размерный и качественный состав пищи зоопланктона фронтальных зон юго-восточной части Тихого океана, мы выделили 4 группы планктеров по доминированию в пище частиц разных размерных категорий (Рис. 1) (Арашкевич, Пастернак, 1984). В группу I (в пище доминирует фракция от 1 до 6 мкм) входят Appendicularia, копеподы родов Paracalanus, Calocalanus, Clausocalanus; в группу II (доминируют частицы 6-40 мкм) - рачки из р.Acrocalanus, Calanus, Neocalanus; в группу III (преобладают пищевые частицы 40-250 мкм) – Eucalanus, Euchirella, Scolecithrix, Centropages, Temora; IV группу составляют животные, потребляющие, главным образом, объекты крупнее 250 мкм, копеподы рода Euchaeta. Эти группы совпадают с принятым ранее разделением планктонных организмов по способу добывания пищи (группы тонких и грубых фильтраторов, животных с комбинированным способом добычи пищи, хватателей) (Тимонин, 1973; Сажин, 1982). Как видно из Рис. 1, границы пищевых спектров полностью перекрываются. Однако пищевые спектры существенно отличаются друг от друга по доминированию в пище той или иной категории частиц. Наиболее «растянутые» спектры, т. е., наибольшая пищевая пластичность у грубых фильтраторов и животных с комбинированным способом добывания пищи.
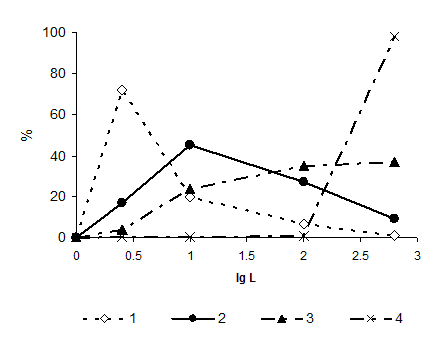
Рис. 1. Размерные пищевые спектры планктеров юго-восточной части Тихого океана. Доля (%) частиц разного размера в пищевом комке копепод. Группа 1: в пище преобладают частицы размеров 1 до 6 мкм; группа 2: преобладают частицы 6-40 мкм; группа 3: преобладают частицы 40-250 мкм; группа 4: преобладают объекты крупнее 250 мкм (по Арашкевич, Пастернак, 1984).
Внутри выделенных трофических групп с увеличением размера консументов увеличивается процентное содержание крупных объектов в их пищевом комке. Крупные консументы при возможности используют более крупные кормовые объекты, сохраняя возможность дополнять свой рацион мелкими. Наши данные согласуются с существующими представлениями об изменениях в структуре ротовых придатков по мере увеличения размера животных (Marshall, 1973; Андронов, 1979; Nival, Nival, 1976; Vanderploeg, Ondricek-Fallscheer, 1982). В конкретной ситуации пищевые спектры, помимо размера консумента и способа добывания им пищи, определяются качественным и размерным составом доступной пищи и соотношением ее компонентов. Рассмотренные особенности пищевых спектров планктеров разных трофических групп хорошо выделяются лишь при разнообразном составе пищи. Так, исследуя питание зоопланктона Тихого океана, мы показали, как сильно могут различаться размерные спектры у представителей одной трофической группы (Арашкевич, Пастернак, 1984) в зависимости от условий питания на разных станциях.
В период максимального обилия фитопланктона все исследованные копеподы потребляли почти исключительно фитопланктон (рис. 2А). Если летом доля неоформленной массы (образующейся при питании инфузориями, голыми жгутиковыми, детритом и бактериальными агрегатами) в кишечниках антарктических копепод составляла около 20% (за исключением Microcalanus pygmaeus), то зимой это - доминирующая составляющая пищевого комка (40-97%, рис. 2 В). Наиболее ярким исключением из этой закономерности служит мелкий рачок Ctenocalanus citer, обитающий и в зимнее время в поверхностных слоях и использующий в пищу ассоциированные со льдом водоросли (Pasternak, Schnack-Schiel, 2007). Крупный Calanoides acutus никогда не содержал пищу зимой.
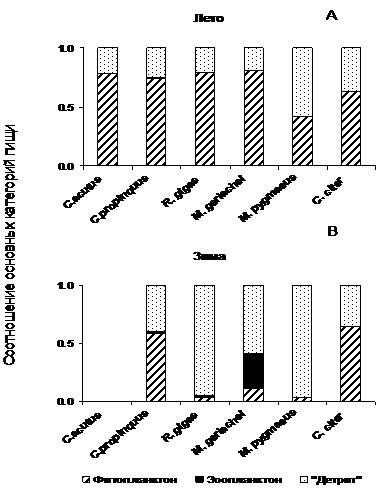

Рис. 2. Основные компоненты пищевого комка (в долях от общего) доминирующих растительноядных копепод (самок и CV) Антарктики летом (A) и зимой (B).
Потенциальные пищевые спектры зависят от морфологии ротовых придатков, размера консумента, сенсорного оснащения копепод, а реализованные - также от концентрации и соотношения различных пищевых объектов в среде. Пищевая пластичность позволяет преимущественно растительноядным копеподам обеспечивать себя кормом в условиях резко меняющейся доступности ресурса. Фитопланктон в период обилия составляет львиную долю их пищи. При уменьшении концентрации фитопланктона одни копеподы продолжают питаться (наногетеротрофами, детритом, ледовыми водорослями, зоопланктоном), а другие полностью прекращают питание, впадая в диапаузу.
3.2. Интенсивность питания. Эмпирическая зависимость рациона от массы тела копепод (Сущеня, 1975) показывает, что с увеличением размера животного его удельный рацион снижается, что справедливо как для межвидовых, так и для внутривидовых сравнений. Мы сопоставили полученные данные по скорости потребления пищи рачками разного размера с общей зависимостью.
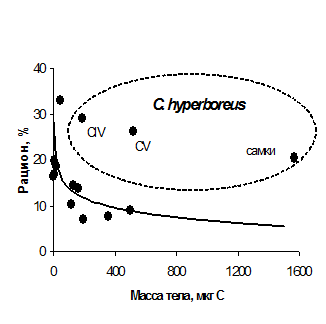
Рис. 3. Зависимость удельного рациона от массы тела массовых видов растительноядных копепод в период цветения в Баренцевом море. Кривая - общая зависимость удельного рациона от массы тела для копепод при избытке пищи (Сущеня, 1975). Точками показаны наши данные для Calanus hyperboreus, C. glacialis, C. finmarchicus, Metridia longa, Pseudocalanus sp., Microcalanus sp., Oithona sp.
Удельный рацион Calanus hyperboreus существенно выше, чем у других, более мелких, копепод, питавшихся растительной пищей в период обилия фитопланктона (Рис. 3). Тем самым нарушается закономерность, согласно которой удельная скорость потребления пищи у крупных потребителей ниже, чем у мелких. В самый ответственный для питания растительноядных копепод период – цветение фитопланктона - самая крупная копепода, C. hyperboreus, демонстрирует необычно высокую скорость потребления пищи.
3.3. Питание в условиях неоднородного размещения ресурсов Чаще всего при исследовании питания в водной среде принимается, что корм размещен случайным образом и частота встреч между потребителем и жертвой определяется средней концентрацией корма, двигательной активностью потребителя и областью его рецепции (Ивлев, 1961; Holling, 1959; Gerritsen, Strickler, 1977). Однако пищевые ресурсы, как правило, распределены неравномерно во времени и пространстве, причем масштабы этой неоднородности различны.
Попытки свести баланс между поступлением энергии при питании планктонного животного и ее расходом на дыхание, рост и размножение часто приводят к выводу о недостатке корма для выживания. Экспериментальные и расчетные данные показывают, что зоопланктону не хватает корма для роста и размножения при средней концентрации фитопланктона, оцениваемой при стандартных сборах (Lasker 1966; Mullin & Brooks 1976; Cowles et al., 1998; Clay et al. 2004). Безусловно, определенную роль играют альтернативные источники пищи, которые не всегда учитываются. Однако чаще всего большую роль играет неоднородность в распределении пищевых частиц.
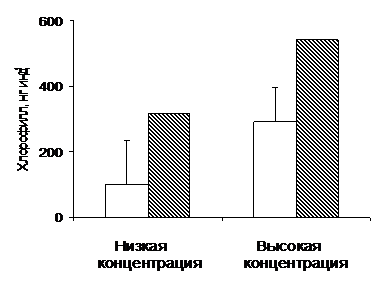 |
Рис. 4. Индексы питания (содержание хлорофилла в кишечнике, индивидуальные оценки) самок Calanus hyperboreus при низкой (0,6-0,8 мкг Chl л-1) и высокой (9-12 мкг Chl л-1) средней концентрации фитопланктона. Светлые столбики - средние значения + SD. Заштрихованные столбики - максимальные значения.
Прямые оценки питания копепод в пятнах и слоях повышенной концентрации, к сожалению, отсутствуют, но полученные нами данные косвенно указывают на эффективное потребление пищи в местах ее скоплений. С помощью флуоресцентной
методики нам удалось получить индивидуальные оценки наполнения кишечника у самок C. hyperboreus на станциях с низкой (0.6-1 мкг хл л-1) и высокой (9-12 мкг хл л-1) концентрациями фитопланктона (Рис. 4). Хотя в среднем C. hyperboreus потреблял намного меньше пищи (в 3 раза) при низкой концентрации, чем при высокой, различия между максимальными величинами потребления были не столь велики. Максимальное потребление пищи при низкой концентрации фитопланктона выше среднего при высокой. Вариабельность в потреблении пищи выше при низком содержании фитопланктона в среде. Это указывает на то, что часть рачков смогла отыскать скопления фитопланктона даже при его низкой, в среднем, концентрации. Поэтому некоторая часть особей имела хорошо наполненные кишечники, а остальные были почти пустыми.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
