
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
В работе основное внимание уделено системным способам защиты. В морской среде у растительноядных копепод широко распространены модификации жизненных циклов, связанные с заблаговременным уходом (в пространстве и во времени) из опасных биотопов. Большинство копепод высоких широт совершает сезонные и онтогенетические миграции, откармливаясь в богатых пищей, но опасных поверхностных слоях летом и погружаясь в глубину зимой. Чаще всего, недостаток корма зимой считался главной причиной ухода из поверхностных слоев.
Начало миграции в глубину у разных видов происходит в разное время. Самки Calanus hyperboreus прекращают питаться и уходят из поверхностных слоев в начале лета, когда концентрация пищи максимальна, а старшие копеподиты (CIV и CV) на месяц позже (Pasternak et al., 2001). C. finmarchicus в это время не только продолжает питаться в верхних слоях, но и демонстрирует максимум пищевой активности. Пресс зрительных хищников на крупных рачков сильнее (Zaret 1980; Гиляров 1987; Lazzaro, 1987), что сдвигает их уход из поверхностных слоев на более раннее время. Копеподы, активные круглый год, обычно обладают мелкими размерами и растянутым размножением (например, Oithona spp., Microcalanus pygmaeus), что позволяет распределить риски (гипотеза «bet hedging») по большей части года. Весь жизненный цикл Ctenocalanus citer проходит в поверхностном слое. Я предполагаю, что сдвиг его максимальной пищевой активности на осенне-зимний период связан с избеганием хищников. Заполненный кишечник увеличивает заметность рачка, поэтому в период максимального обилия хищников в весенне-летний период он снижает питание.
Изменения в жизненном цикле под влиянием хищников возникают и у пресноводных копепод, для которых механизмы взаимодействия удалось исследовать более подробно. Нами была обнаружена «личиночная диапауза» (sensu Алексеев, 1990) в летнее время у копеподы Eudiaptomus graciloides (Pasternak et al., 1996; Pasternak, Arashkevich, 1999) в озере с высоким прессом рыб-планктофагов (Boikova, 1986). Вместо сменяющих друг друга пиков последовательных возрастных стадий обнаружена аккумуляция старших копеподитов (Рис. 11), большая часть которых погружалась в
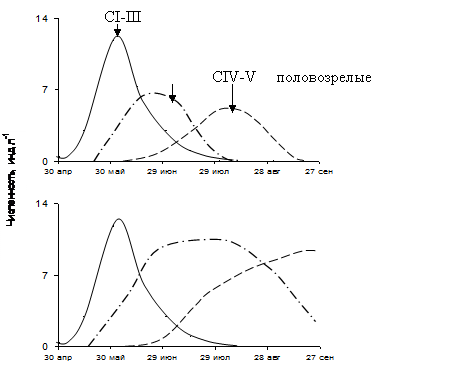 |
Рис. 11. Различные стратегии жизненного цикла Eudiaptomus graciloides. Вверху: типичное чередование последовательных копеподитных стадий при низком прессе хищников (оз. Кривое, по , 1975). Внизу: аккумуляция старших стадий из-за летней диапаузы при высоком прессе хищников в оз. Глубоком (по Pasternak, Arashkevich, 1999).
гиполимнион. Скорость развития (с поправкой на температуру) у особей в гиполимнионе, где они были недоступны для хищников, снижалась более чем в 2 раза. В озере Кривом, где пресс планктофагов был невелик, популяция E. graciloides развивалась непрерывно в течение летнее-весеннего сезона (Иванова, 1975). Diaptomus sanguineus переходил от продукции субитанных к откладке покоящихся яиц в прудах с высокой численностью рыб-планктофагов (Hairston 1987).
5.2. Ответные способы.
При возникновении угрозы хищничества окрашенные рачки становятся более прозрачными (Hansson, 2004; Vestheim, Kaartvedt, 2006), что может рассматриваться как «ответный» механизм защиты. Широко распространенная тактика защиты зоопланктона от хищников, суточные вертикальные миграции (СВМ), проявляется по-разному при разном прессе хищников. Так, увеличение концентрации кайромонов хищников (рыб и гребневиков) меняло характер и параметры СВМ у Calanopia americana (Cohen, Forward, 2005). Мы показали, что при увеличении пресса рыб-планктофагов у Eudiaptomus gracilis возникают миграции, приводящие к образованию плотных придонных скоплений рачков в сублиторали.
В рамках данной работы наибольшее внимание привлекают системные изменения (изменения в жизненных циклах), происходящие в ответ на появление хищников. К системным изменениям относятся более ранее генеративное созревание при меньших размерах или увеличение размера кладки, что наблюдали у кладоцер (Stibor, 1992; De Meester et al., 1995; Gliwicz, Boavida, 1996; Sakwińska, Dawidowicz, 2005). Хотя прямых сведений о подобных модификациях цикла у копепод к настоящему времени нет, некоторые результаты позволяют предположить, что сходные ответы могут быть и у морских копепод. Так, размер половозрелых самок копепод внутри популяции может различаться на 20-30% (например, по нашим данным, самки зимней и весенней генерации C. finmarchicus из Норвежского моря). При угрозе самки циклопов могут раньше приступать к формированию яйцевых мешков, чем в благоприятной ситуации.
х х х
Мы предлагаем следующую схему основных оборонительных тактик растительноядных копепод (рис. 12). Граница между упреждающими и ответными способами защиты условна, однако в своих крайних проявлениях они хорошо различаются. При угрозе, исходящей от рыб-планктофагов, разница в размерах и скорости плавания между хищником и жертвой настолько значительна, что ответные реакции гораздо менее эффективны, чем упреждающие. Предотвратить возможную встречу с хищником, заблаговременно покинув опасный биотоп, более выгодно. 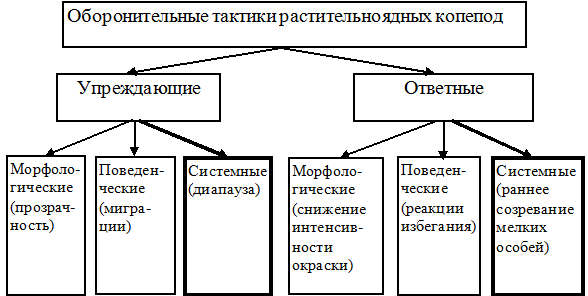
Рис. 12. Оборонительные тактики растительноядных копепод (в скобах даны примеры).
На доступность копепод хищникам может существенно влиять зараженность паразитами, которая влияет на все аспекты экологии хозяев и модифицирует их жизненные циклы. Данных для морских копепод к настоящему времени мало для того, чтобы уверенно оценивать роль паразитов в жизненных циклах рачков. Хищничество и паразитизм, будучи мощными факторами отбора, часто действуют совместно.
Глава 6. Диапауза и переживание неблагоприятных условий: экологические и метаболические аспекты.
Наиболее радикальной из адаптаций копепод, способствующих переживанию неблагоприятных периодов, является диапауза. Это состояние у копепод аналогично диапаузе насекомых (Elgmork, Nilssen, 1978). Помимо защитной функции, диапауза играет важную роль, синхронизируя размножение и рост младших стадий в благоприятных условиях (Santer, Lampert, 1995). Диапауза у копепод наблюдается на разных стадиях развития и подразделяется на эмбриональную, личиночную и имагинальную (Алексеев, 1990; Zeller et al., 2004). У животных в диапаузе снижается метаболизм и подвижность, приостановливается рост и развитие (Алексеев, 1990; Hutchinson, 1967; Mauchline, 1998).
Миграции – уход от неблагоприятных условий в пространстве, диапауза - во времени. В морях диапауза тесно связана с перемещением в пространстве. При эмбриональной диапаузе, характерной для прибрежных и мелководных районов, покоящиеся яйца копепод погружаются на дно. Старшие копеподиты и взрослые особи впадают в диапаузу, опускаясь в глубину.
6.1. Накопление и расходование липидов. Растительноядные копеподы в период цветения фитопланктона накапливают липиды, позволяющие пережить зиму без пищи в диапаузе, а следующей весной перелинять, подняться к поверхности и приступить к размножению. Высокое содержание энергии делает липиды наиболее подходящим энергетическим резервом (Lee et al., 2006).
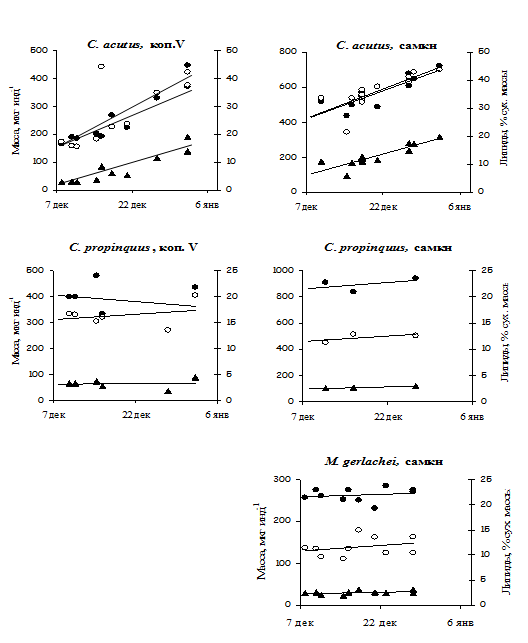 Мы полагаем, что два периода в основном определяют динамику жировых резервов: цветение фитопланктона, во время которого идет накопление липидов, и переход от диапаузы к активной жизнедеятельности, во время которого основная часть липидов утилизируется. На квазистационарной станции в море Уэдделла во время развития весеннего «цветения» нам удалось детально проследить динамику накопления липидов у копепод с разным способом переживания неблагоприятных условий. Общее содержание липидов у CV C. acutus за три недели увеличилось более чем в 2 раза, а у Metridia gerlachei и Calanus propinquus лишь на 1-4% (Рис. 13). Скорость накопления
Мы полагаем, что два периода в основном определяют динамику жировых резервов: цветение фитопланктона, во время которого идет накопление липидов, и переход от диапаузы к активной жизнедеятельности, во время которого основная часть липидов утилизируется. На квазистационарной станции в море Уэдделла во время развития весеннего «цветения» нам удалось детально проследить динамику накопления липидов у копепод с разным способом переживания неблагоприятных условий. Общее содержание липидов у CV C. acutus за три недели увеличилось более чем в 2 раза, а у Metridia gerlachei и Calanus propinquus лишь на 1-4% (Рис. 13). Скорость накопления
|
липидов в период цветения у C. acutus выше, чем у других антарктических видов. Удельный рацион C. acutus выше, чем у C. propinquus. У экологически сходного арктического вида С. hyperboreus удельная скорость питания в период цветения такжн значительно выше, чем у более мелких совместно обитающих копепод (Рис. 3). Оба периода, накопления и расходования липидов, были охвачены круглогодичными измерениями объема жирового мешка у CV С. hyperboreus и C. finmarchicus, которые мы проводили во фьорде Маланген в северной Норвегии. Объем жира у С. hyperboreus начал увеличиваться в марте, когда фитопланктон лишь начал развиваться, а в апреле уже достиг максимума (Рис. 14), в то время как у C. finmarchicus жир накапливался постепенно с мая (максимум цветения) по август (Pasternak et al., 2001). У С. hyperboreus, вида с более продолжительной диапаузой, как питание, так и накопление жира происходит с чрезвычайно высокой интенсивностью в очень ограниченный период.
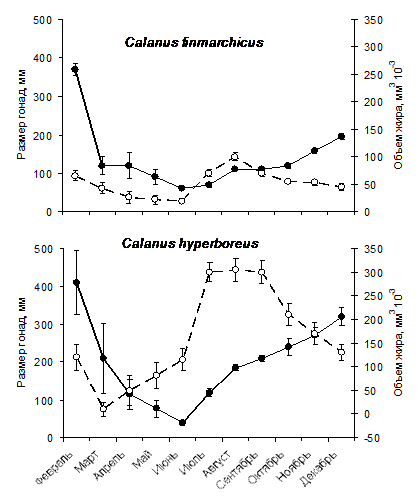 |
Рис. 14. Размер гонад (сплошная линия) и объем жирового мешка (прерывистая линия) у CV Calanus finmarchicus и C. hyperboreus из фьорда в северной Норвегии (слой 100-200 м). С конца лета на фоне уменьшения объема жира увеличивается размер гонад.
У растительноядных копепод высоких широт основным резервным веществом служат 2 класса липидов: восковые эфиры (ВЭ) и триацилглицерины (ТАГ). У видов с продолжительной диапаузой содержание ВЭ обычно высоко, за исключением C. propinquus, у которого преобладают ТАГ (Schnack-Schiel et al., 1991; Hagen et al., 1993; Kattner, Hagen, 1995). Наибольшие запасы высококалорийных ВЭ обнаружены у видов с наиболее продолжительной (до 8-9 месяцев) облигатной диапаузой, C. hyperboreus в северном полушарии и C. acutus в южном. Мы показали, что жировые запасы самок C. acutus в декабре на 85-90% состоят из ВЭ, доля ТАГ не превышает 1%. Содержание ВЭ у CIV и CV в среднем ниже (соответственно 56 и 78%). Доля жирных кислот и спиртов с 22 атомами углерода, обеспечивающих повышенную калорийность жира, у этого вида также значительно выше, чем у остальных антарктических видов. Содержание ВЭ в начальный период цветения у C. propinquus составляло 0.4-2%, у M. gerlachei - 7%, а ТАГ – 35-55% и 10%, соответственно.
По нашим расчетам, энергия накопленных липидов во много раз превышает расходы на базовый метаболизм C. propinquus (Drits et al., 1993). Это согласуется с современными представлениями о расходовании запасенных липидов в основном не на метаболизм, а на развитие гонад и ооцитов, а также подъем к поверхности (Hirche, Kattner, 1993; Hagen, Schnack-Schiel, 1996; Jónasdóttir, 1999). Одновременная оценка объема жира и размера гонад у CV С. hyperboreus и C. finmarchicus выявила их реципрокную связь (Pasternak et al., 2001). Та же зависимость выявлена нами и для самок - зрелые самки C. acutus содержали меньше жира с более низким содержанием ВЭ.
Как общий запас жира, так и его состав указывают на то, что к длительной зимовке без пищи наиболее приспособлены C. hyperboreus на севере и C. acutus на юге. Скорость накопления и общий запас липидов у них выше, чем у других видов. Выполненные нами одновременные исследования жировых запасов и стадии зрелости гонад арктических и антарктических копепод подтвердили представление о важнейшей роли липидов в генеративном развитии копепод.
6.2. Снижение уровня метаболизма. Нами впервые были получены оценки скорости метаболизма в «глубинной» и «поверхностной» частях популяций трех видов антарктических копепод. Скорость дыхания, определенная по активности ЭТС, у погрузившихся самок и CV Calanoides acutus из моря Уэдделла была, с поправкой на температуру, в 8 раз ниже, чем у остававшихся в поверхностном слое особей. Экскреция аммония и фосфата снижалась более чем вдвое (Рис. 15). Также пониженная скорость экскреции и дыхания наблюдалась у «глубинных» Calanus propinquus, в то время как дыхание самок и CV Rhincalanus gigas в поверхностных и глубинных слоях не различалось (Pasternak et al., 1994; Pasternak, Schnack-Schiel, 2001). Это показывает, что погрузившиеся C. acutus и C. propinquus были в состоянии диапаузы, а R. gigas - в активном состоянии.
|

Рис. 15. Скорость дыхания и экскреции у массовых видов антарктических копепод из поверхностных (0-100 м, светлые прямоугольники) и глубинных ( м, заштрихованные прямоугольники) слоев в период перехода от активной фазы развития популяций к диапаузе. Данные приведены к средней температуре поверхностного слоя.
В северном полушарии у зимующего Calanus hyperboreus дыхание снижалось
в 2 раза (Auel et al., 2003), а у C. finmarchicus в 5 раз по сравнению с активными особями в поверхностном слое (Hirche, 1983, 1996). Практическая неподвижность животных в диапаузе делает их менее заметными для хищников, особенно «тактильных» (Gerritsen, Strickler, 1977; Greene, 1986; Bagøien et al., 2001), а низкий уровень метаболизма позволяет экономить энергию, необходимую для зимовки, развития гонад, линьки и подъема к поверхности после окончания диапаузы.
6.3. Развитие генеративной системы во время диапаузы. Остановка роста и развития копепод считается одним из основных признаков диапаузы. Тем не менее, в период диапаузы происходит закладка и, по меньшей мере, начальный этап созревания ооцитов. Таким образом, диапауза не означает полной остановки развития, так как генеративная система продолжает развиваться. Кроме того, часть энергии тратится на линьку.
Так как в диапаузе копеподы не питаются, жизнедеятельность и развитие генеративной системы обеспечиваются накопленным жиром. Примерно половина жировых запасов Calanus helgolandicus расходуется на гонадогенез и линьку CV в половозрелых особей (Gatten et al., 1980). Однако было неизвестно, происходит ли развитие и рост гонад только после того, как пройдена большая часть диапаузы и внутренние механизмы сигнализируют, что жира сохранилось достаточно для завершающего этапа, или развитие гонад идет в течение всей диапаузы.
Чтобы выявить характер сезонного развития гонад и динамики липидов, мы оценивали состояние гонад и объем жира у CV Calanus hyperboreus и C. finmarchicus в течение всего года. Полученные результаты показали, что в течение всей диапаузы (с июля по декабрь у C. hyperboreus и с августа по февраль у C. finmarchicus) происходит постепенное увеличение в размере и дифференциация гонад у особей в глубинных слоях (Рис. 14, Pasternak et al., 2001). Параллельно у обеих копепод уменьшается объем жира. В течение всего периода, но чаще к окончанию диапаузы в популяциях встречаются особи, не содержавшие липидов, и, по-видимому, обреченные на вымирание. У многих из них гонады были хорошо развиты. Их дальнейшее развитие, очевидно, останавливалось лишь нехваткой энергии. Созревание гонад у CV C. hyperboreus начиналось и завершалось раньше, чем у совместно обитающего C. finmarchicus, что согласуется с ранним началом размножения первого вида.
Полученные результаты убедительно свидетельствуют, что в период диапаузы у CV происходит непрерывное развитие гонад параллельно с расходованием жировых запасов, линька в половозрелых особей и созревание ооцитов.
6.4. Сигналы к началу и окончанию диапаузы. Стимулы к началу и завершению диапаузы, в основном изучавшиеся на пресноводных копеподах (обзоры – Алексеев, 1990; Dahms, 1995; Williams-Hoze, 1997), до конца не выяснены. Как возможные триггеры рассматривались изменения освещенности, температуры, концентрации пищи, уровня жиронакопления и пресса хищников (Cooley, 1978; Алексеев,1984, 1990; Marcus, 1980; Hairston, 1987; Kaartvedt, 1996; Pasternak, Arashkevich, 1999; Niehoff, Hirche, 2005). Большинство авторов склоняется к тому, что основной сигнальный фактор – фотопериод, а температура лишь модифицирует его эффект (Hairston et al., 1990; Ban, 1992; Pasternak, Arashkevich, 1999). В морских биотопах освещенность также рассматривалась как фактор, инициирующий погружение в глубину и впадение в диапаузу у каляноид (на примере Calanus finmarchicus, Miller et al., 1991). Другие авторы нашли, что C. glacialis прекращает питаться и размножаться при повышении температуры в поверхностном слое выше 6оС и уходит в диапаузу, несмотря на высокую концентрацию корма в поверхностных слоях (Kosobokova, 1999; Niehoff, Hirche, 2005).
По нашим данным, длина фотопериода играла решающую роль при переходе Eudiaptomus graciloides от продукции субитанных к откладке покоящихся яиц. Увеличивая в экспериментальных условиях осенью длину фотопериода, мы смогли задержать переход к продукции покоящихся яиц самками E. graciloides (Pasternak, Arashkevich, 1999). Изменяя температурные и пищевые условия, мы не получили подобного результата. Сигналы к терминации диапаузы мы исследовали, помещая покоящиеся яйца в темноту при пониженной температуре (7о С) на 2-3,5 месяца. Науплии появлялись после перенесения покоящихся яиц в условия повышенной температуры (20-21о С). Вылупление происходило даже в темноте, но ускорялось на свету. Науплии выклевывались более дружно при имитации зимних условий в течение 3,5, чем 2 месяцев. Эти результаты хорошо согласуются с данными по насекомым, где повышение температуры до завершения эндогенной фазы (наиболее глубокая диапауза, связанная с относительной независимостью от внешних условий) приводит к десинхронизации процесса активации (Mansingh, 1971; Алексеев, 1990). По завершении эндогенной фазы животные становятся восприимчивы к внешним сигналам.
Мы полагаем, что основным сигнальным фактором, инициирующим вхождение копепод в диапаузу, служит изменение фотопериода, как наиболее регулярного явления, надежно указывающего на изменения в среде. Вопрос о терминации диапаузы сложнее, так как трудно согласиться с мнением, что на глубине около 1000 м животные оценивают изменения в освещенности (Miller et al., 1991). Внутренние «биологические часы» или спонтанная реактивация представляются более вероятными.
6.5. Диапауза и миграции. Практически все крупные массовые виды копепод-фитофагов из высоких широт обитают в большом диапазоне глубин и зимой совершают миграции из поверхностных слоев на глубину (Виноградов, 1968; Воронина, 1984). Глубина погружения и рассредоточение в столбе воды или, напротив, концентрация в сравнительно узком слое различаются. Глубже всего (до м у Calanus hyperboreus и Calanoides acutus) обычно погружаются копеподы в состоянии диапаузы. Однако не все копеподы, опустившиеся зимой в глубину, находятся в диапаузе. Metridia gerlachei, погрузившись зимой в глубину, переходит к хищничеству и/или детритофагии, сохраняя активность весь год. У более мелких копепод (Microcalanus pygmaeus, Ctenocalanus citer), остающихся активными зимой, амплитуда сезонных миграций меньше. Крупные Calanus propinquus и Rhincalanus gigas, погружаясь в глубину зимой, в зависимости от условий питания могут впадать в диапаузу или оставаться активны. По сравнению с облигатно диапаузирующим C. acutus, эти копеподы могут питаться более крупными пищевыми частицами и хищничать, а их липидные резервы ниже. Диапауза у них не такая глубокая, и при попадании в благоприятные условия они могут сравнительно быстро перейти в активное состояние.
Так как в диапаузе копеподы малоподвижны и не реагируют на внешние стимулы, в верхних слоях они были бы беззащитны перед зрительными хищниками. Лишь уходя в недоступные для зрительных хищников (рыб) слои, рачки обеспечивают себе относительную безопасность. Копеподам, сохраняющим активность зимой, выгодно рассредоточиться – это позволяет им снизить риск выедания хищниками и конкуренцию за скудный пищевой ресурс. Это подтверждается их более «растянутым» профилем вертикального распределения зимой.
х х х
Возможность использования такого мощного защитного и синхронизирующего механизма, как диапауза, крайне важна при переживании неблагоприятного периода в среде с выраженной цикличностью. У морских копепод диапауза обязательно сопровождается погружением в глубокие слои воды. Возможность впадать в диапаузу зависит от способности копепод накапливать значительные липидные резервы, в первую очередь, высококалорийные восковые эфиры. Необходимая для этого быстрая аккумуляция энергии обеспечивается интенсивным питанием в плотных скоплениях корма. Принципиальным является выход из диапаузы и начало активной фазы к началу «цветения» фитопланктона.
Глава 7. Стратегии жизненных циклов растительноядных копепод в среде с резко выраженной цикличностью.
Обычно питание, размножение, диапауза и особенности жиронакопления копепод высоких широт рассматривались по отдельности, чаще всего в весенне-летний период. Основное внимание уделялось закономерностям питания, и циклический характер обилия пищи помещался во главу угла при объяснении особенностей жизненного цикла (например, Hirche, 1996). В дальнейшем больше внимания стали уделять генеративным стратегиям (Hirche, Kosobokova, 2003; Pasternak et al., 2004 и др.). Роль такого важного экологического фактора, как пресс хищников, в формировании жизненных циклов копепод, недооценена. Мы полагаем, что сезонный пик пресса хищников может оказывать не меньшее влияние на уход копепод в диапаузу, чем снижение концентрации пищи.
7.1. Основные типы жизненных циклов. Я рассматриваю формирование жизненного цикла растительноядных копепод как компромисс между требованиями питания, размножения и защиты от хищников. Присущие разным копеподам характерные особенности этих процессов и их различные сочетания приводят к возникновению многообразия жизненных циклов.
Предлагаемая концепция позволила свести это многообразие к четырем основным типам (рис. 16). Прежде всего, это 2 типа циклов, связанных с избеганием «трофогенной зоны» в неблагоприятный сезон и 2 типа циклов, связанных с приспособлением к ней. Под «трофогенной зоной» мы понимаем верхние слои воды с повышенной концентрацией доступной пищи и зрительных хищников. Копеподы, использующие стратегии избегания, уходят на глубину и впадают в диапаузу при сезонном уменьшении доступности пищи.
 |
Рис. 16. Стратегии жизненных циклов растительноядных копепод высоких широт. Репродуктивный запас энергии – часть общего запаса энергии, необходимая для размножения.
Для таких стратегий характерны: короткий (весна-лето) период размножения и накопление в продуктивный сезон значительных липидных резервов, в основном, восковых эфиров. Копеподы со стратегией приспособления не уходят в диапаузу, а остаются активными круглый год. Для этих стратегий характерны: растянутый период размножения и значительно меньшие липидные резервы (в основном, триацилглицерины), которые служат не для переживания продолжительного неблагоприятного периода, а для подстраховки при локальном дефиците корма.
Далее разделение жизненных циклов идет по тому, как пополняется запас энергии, обеспечивающей размножение. Среди планктеров, использующих диапаузу, выделяется группа, накапливающая энергию лишь в сравнительно короткий период, предшествующий диапаузе; их размножение происходит целиком за счет резервных липидов. При этой стратегии периоды размножения и питания разнесены во времени. Характерный пример – Calanus hyperboreus. Необычайно высокая интенсивность питания в продуктивный период позволяет ему быстро накопить значительные жировые запасы. Очень короткий период активности связан, по-видимому, с сезонным пиком зрительных хищников в поверхностном слое. Раннее размножение обеспечивает быстрое развитие потомков и их заблаговременный (до пика зрительных хищников) уход в глубину. В исследовании на модели жизненного цикла Calanoides acutus было показано, что в наиболее благоприятном положении оказалось потомство, полученное из самых ранних яиц (Varpe et al., 2007). Хотя численность такого «раннего» потомства часто невысока, его роль в успешном развитии популяций непропорционально высока (Smith, 1990; Varpe et al., 2007).
Другая группа объединяет циклы, в которых размножение, хотя и происходит в значительной мере за счет липидов, начинается лишь при поступлении энергии с началом питания в новом сезоне. Характерный пример - C. finmarchicus. При этом периоды размножения и питания не разнесены во времени, как в описанной выше стратегии.
Копеподы со стратегиями приспособления непрерывно пополняют запас энергии. Растянутый период питания и размножения позволяет им распределить риски по разным сезонам и стадиям развития популяции (bet-hedging). Среди этих копепод одна группа питается в зимний период на «концентрированном ресурсе», т. е., использует локальные пятна фитопланктона и наногетеротрофов, слои повышенной концентрации взвеси. Эти копеподы отличаются практически круглогодичным размножением и относительно мелкими размерами. Характерный пример - Microcalanus pygmaeus, Ctenocalanus citer. Другая группа в зимний период использует «рассредоточенный ресурс», переходя к преимущественно хищному питанию и потребляя более крупные и питательные, хотя и более редкие, частицы. Характерный пример - Metridia gerlachei, M. longa. При использовании рассредоточенного ресурса и потребители его рассредоточены, что, в свою очередь, снижает риск быть съеденными хищниками.
7.2. Роль размеров копепод. Параметры жизненных циклов взаимосвязаны и наличие одних черт лимитирует диапазон вариаций других в пределах возможностей организма. В экологических исследованиях большое внимание уделяется размерам организмов (Schmidt-Nielsen, 1984; Begon et al., 1990 и др.). Скорость основных процессов жизнедеятельности ракообразных зависит от их размеров (Винберг 1950; Сущеня, 1975). Размеры копепод играют важнейшую роль в формировании их жизненных стратегий, создавая для крупных организмов предпосылки для диапаузы, мелких же копепод в значительной степени выводя из-под пресса рыб-планктофагов.
Мелкие организмы тратят больше энергии на единицу массы тела, чем их более крупные сородичи, поэтому для мелких рачков любые миграции, в том числе, связанные с диапаузой, обходятся дороже. Удельные траты энергии на дыхание у мелких копепод выше, чем у крупных. Даже при равных удельных запасах липидов (а они выше у крупных копепод, см. Lee et al., 2006), этих резервов крупным копеподам будет хватать на более продолжительную зимовку. Число крупных видов с достоверно описанной диапаузой существенно выше, чем мелких. Более того, мы показали, что виды мелких копепод с предполагавшейся ранее диапаузой (Microcalanus pygmaeus, Ctenocalanus citer - Atkinson, 1998) были активны весь год (Pasternak, Schnack-Schiel, 2001, 2007).
Стратегии питания, размножения и защиты от хищников также связаны с размером копепод. Ранее мы показали, что с увеличением размера потребителя растет максимальный размер доступных ему жертв. В рационе крупных копепод увеличивается доля крупной добычи, хотя сохраняется способность потреблять и мелкие частицы, поэтому размерный диапазон пищевых частиц у крупных рачков шире. Кроме того, им легче справиться с подвижной добычей. Но концентрация пищи, при которой происходит насыщение, для них выше, чем для мелких рачков (Nival, Nival, 1976), поэтому при низких концентрациях мелкие рачки получают преимущество. Этим можно объяснить высокий процент мелких рачков среди активно питающихся зимой копепод.
Количество откладываемых яиц зависит от размера самки – более крупные самки, содержащие больше липидов, единовременно производят больше яиц (Hirche, 1989; Smith, 1990; Runge, 1984; Pasternak et al., 2004). С другой стороны, период репродукции обычно более продолжителен у мелких копепод. Тем самым мелкие рачки увеличивают общее число производимых потомков и распределяют риски во времени.
Зрительные хищники при прочих равных условиях избирают более крупных жертв, (Ивлев, 1955; Zaret, 1980). Это может определять ранний (в период, когда корма наверху еще много), уход крупных копепод, размножающихся до и во время пика цветения, из трофогенной зоны в глубину. Мелкие рачки менее уязвимы для рыб-планктофагов, что позволяет им питаться и размножаться практически круглый год. Уменьшение размера, при котором достигается половозрелость, еще один способ снизить угрозу хищничества (Stibor, Lampert, 2000; Sakwinska, 2002; Thys, Hoffmann, 2005).
В предлагаемой классификации жизненных циклов (Рис. 16) мелкие копеподы (Microcalanus pygmaeus, Ctenocalanus citer, Oithona similis), а также несколько более крупные Metridia gerlachei, M. longa располагаются в правой части схемы (циклы без диапаузы, с незначительными жировыми резервами, непрерывным питанием и размножением). Наиболее крупные рачки (Calanus hyperboreus, C. glacialis, Calanoides acutus) занимают левую часть схемы, придерживаясь стратегии с уходом в продолжительную диапаузу, накоплением больших липидных резервов и коротким периодом пищевой и репродуктивной активности. Таким образом, размер копепод определяет вероятность, с которой они будут следовать той или иной стратегии жизненного цикла. У крупных копепод больше возможностей выбора между стратегиями, чем у мелких.
7.3. Вариабельность жизненных циклов. Стратегия жизненного цикла – это комплекс поведенческих и физиологических реакций животного, направленных на разрешение основных жизненных задач, и в конечном итоге, на повышение приспособленности. Выбор стратегии происходит на уровне индивидуума и в пределах нормы реакции находится под контролем генотипа (Stearns, 1992; Charnov, 1993).
Далеко не всегда наблюдаемая изменчивость в жизненно важных характеристиках рачков говорит о различиях в стратегиях жизненных циклов. Параметры жизненного цикла в пределах одной и той же стратегии могут модифицироваться конкретными условиями обитания. Региональная изменчивость может быть связана с местными условиями освещенности, временем наступления и продолжительностью цветения фитопланктона. Так, крупные антарктические копеподы Calanoides acutus, Calanus propinquus и Rhincalanus gigas поднимаются к поверхности раньше и остаются там дольше в северной зоне Южного океана, чем в южной (Воронина, 1969, 1974; Voronina, 1970). Время появления новой генерации запаздывает в южном направлении, но от этой тенденции имеется ряд локальных отклонений в сторону более раннего развития в зонах дивергенций, в заливах, рано освобождающихся ото льда (Воронина, 1975, 1984).
В других случаях различия в параметрах жизненного цикла у разных популяций вида или в пределах одной популяции столь велики, что позволяют говорить о разных стратегиях. Мы обнаружили внутрипопуляционную неоднородность по важнейшему признаку, переживанию неблагоприятных условий в пассивном (диапауза) или активном состоянии, у C. propinquus и R. gigas (Pasternak, Schnack-Schiel, 2001). Большая часть рачков в популяциях этих копепод в море Уэдделла находилась зимой в диапаузе, но от 5 до 15% сохраняли активность. Разница во мнениях исследователей на наличие диапаузы у этих копепод (Marin, 1988; Schnack-Schiel, Hagen, 1994; Ward et al., 1997; Pasternak, Schnack-Schiel, 2001) связана с использованием осредненных показателей, характеризующих популяцию (вид). При использовании индивидуумами в популяции различных стратегии результат осреднения будет зависеть от соотношения в выборке особей с той или иной стратегией. В наиболее контрастных биотопах (например, в северных и южных областях ареала) могут преобладать (вплоть до 100%) особи с той или иной стратегией. В ситуациях не столь контрастных соотношение особей с разными стратегиями может варьировать (например, 60:40 или 75:25), что при осредненных показателях может привести к мысли о промежуточных стратегиях. Если измерения и анализ проводятся на уровне особей, которые и принимают жизненно важные решения, то таких противоречий во взглядах можно избежать.
Между видом копепод и стратегией их жизненного цикла нет однозначного соответствия. У одного и того же вида часть особей может впадать в диапаузу, а другая - нет (C. propinquus), часть особей может размножаться до начала питания, а часть – только начав питание (C. glacialis). То есть, у одного вида можно наблюдать как одну, так и несколько стратегий жизненного цикла.
7.4. Разнообразие классификаций жизненных циклов. В одной из первых работ, анализирующих жизненные циклы копепод из высоких широт на основе структурных и эколого-физиологических показателей, авторы (Conover, Huntley, 1991) разделяли копепод в первую очередь по трофическому положению. Обилие фитопланктона полагалось определяющим фактором. Такой подход привел к ошибочному выводу о сходстве жизненных циклов у Calanus hyperboreus и Rhincalanus gigas. Объяснение причин раннего (еще во время «цветения») прекращения питания и ухода в глубинные слои C. hyperboreus и C. glacialis необходимостью приурочить развитие потомства к «цветению» неубедительно. Авторы не придают значения тому, что крупные и накопившие липиды копеподы становятся хорошо заметны для рыб-планктофагов, численность которых к этому времени существенно возрастает. Поэтому, на наш взгляд, наиболее привлекательные для планктоядных рыб крупные копеподы покидают поверхностный слой раньше, чем мелкие.
В других обобщениях по стратегиям жизненных циклов антарктических копепод авторы (Atkinson, 1998; Schnack-Schiel, 2001), также используя как структурные, так и эколого-физиологические данные, пришли к несколько различающимся выводам. Шнак-Шиль отмечает 2 стратегии: одна – с онтогенетическими миграциями и диапаузой, другая – без диапаузы. Первой стратегии придерживается лишь Calanoides acutus, в то время как большинство антарктических копепод не имеют покоящихся стадий. Аткинсон выделил 3 основные стратегии: 1) питание фитопланктоном летом, короткий период размножения, за которым следует диапауза на глубине (C. acutus, Ctenocalanus citer); 2) всеядные – детритоядные рачки, растянутый период питания, роста и размножения и менее выраженная диапауза в глубине (Metridia gerlachei, Calanus propinquus, Oithona similis, Oncaea spp., Microcalanus pygmaeus,); 3) питание, связанное зимой с морским льдом (науплии и копеподиты Stephos longipes и Paralabidocera antarctica). Автор полагает, что Rhincalanus gigas занимает промежуточную позицию между первой и второй стратегией. Ни тот, ни другой автор прямо не определили основные критерии, по которым проводится выделение стратегий, но, судя по сделанным ими выводам, это – отношение к диапаузе (Schnack-Schiel, 2001) и отношение к диапаузе плюс основной объект питания (Atkinson, 1998). Однако при таком разделении возникает ряд сложностей. Прежде всего, авторы не поясняют, на основании каких характеристик, популяционных или индивидуальных, проводится выделение стратегии. Как результат, появляется представление о промежуточных стратегиях, число которых может возрастать при увеличении объемов информации. Вторая проблема – основной объект питания. Показано, что спектры питания большинства этих копепод перекрываются. Если говорить о питании летом во время «цветения» фитопланктона, то все они потребляют почти исключительно фитопланктон. Различия проявляются в осенне-зимне-весенний период при снижении концентрации доступного корма. Однако M. gerlachei, C. propinquus, O. similis, M. pygmaeus, отнесенные Аткинсоном ко 2-ой группе, различаются между собой в это время не меньше, чем C. acutus (1-ая группа) от C. propinquus.
Из расхождений по более конкретным вопросам следует отметить, что по нашим данным, C. citer и M. pygmaeus совсем не включают диапаузу в свой цикл (а не «обладают менее выраженной диапаузой, как полагал Аткинсон). Эти виды сохраняют пищевую активность круглогодично и даже подо льдом (Pasternak, Schnack-Schiel, 2001, 2007)..
х х х
Предлагаемый нами подход позволяет трактовать формирование стратегий жизненных циклов на основе компромиссного распределения доступных ресурсов. Разные варианты разрешения этого компромисса приводят к возникновению различных стратегий или типов жизненных циклов. Разработанная концептуальная схема позволяет проанализировать взаимосвязи между основными процессами жизнедеятельности. Выявленные корреляции позволяют считать, что эта схема обладает определенной прогностической ценностью. Если исследуемый рачок обладает крупным размером, высока вероятность того, что он быстро накопит липидные резервы и заблаговременно покинет биотоп с высоким прессом хищников. Наличие каких-либо двух признаков связано со вполне определенным набором других. Например, ярко выраженное предпочтение растительной пищи и способность накапливать значительные липидные резервы с высокой вероятностью связаны с продолжительным нахождением в диапаузе и ограниченным периодом размножения. Использованный подход позволяет выявить наиболее вероятные сочетания жизненно важных параметров и выявить основные типы жизненных циклов.
Выводы.
1. Жизненные циклы копепод формируются на основе компромисса между требованиями питания, размножения и защиты от хищников. Этот подход позволил свести многообразие жизненных циклов копепод высоких широт к четырем основным типам. Прежде всего, это циклы с диапаузой и без нее. Среди первых выделены стратегия, в которой периоды размножения и питания разделены во времени (Calanus hyperboreus), и стратегия, в которой размножение происходит одновременно с питанием (C. finmarchicus). Среди вторых выделена стратегия с круглогодичным размножением и питанием на концентрированном ресурсе (слои повышенной концентрации мелких частиц) зимой (Microcalanus pygmaeus), и стратегия с ограниченным периодом размножения и зимой преимущественно хищным питанием (Metridia gerlachei).
2. Выделены 4 типа пищевой активности на основе приуроченности времени начала питания к «цветению» фитопланктона и продолжительности периода питания, пищевой пластичности и пищевым предпочтениям при низкой концентрации корма. Тип I (Calanus hyperboreus) – раннее (до «цветения» фитопланктона) начало короткого периода питания. Тип II (C. finmarchicus) – питание начинается в период «цветения». При выраженном сезонном пике питание с более низкой интенсивностью происходит в течение большей части года. Тип III (Metridia gerlachei) - нет перерыва в питании, и пищевая активность в осеннее-зимний период снижается незначительно. Тип IV (Ctenocalanus citer) – питание происходит в течение всего года с максимумом в осенне-зимний период.
3. Установлены основные типы репродуктивных стратегий копепод высоких широт: I - размножение с резким сезонным пиком до «цветения» фитопланктона, обеспеченное липидными резервами (Calanus hyperboreus). II - размножение с выраженным пиком в период цветения фитопланктона (C. finmarchicus, C. propinquus, Rhincalanus gigas). Копеподы с этим типом репродуктивной стратегии размножаются после начала питания, используя как энергию липидов, так и вновь потребленной пищи. III – растянутое размножение, слабо связанное с цветением фитопланктона (Microcalanus pygmaeus, Metridia gerlachei, M. longa, Ctenocalanus citer). Эти копеподы размножаются и питаются в течение всего года
4. Впервые показано, что рост и дифференциация гонад у Calanus finmarchicus и C. hyperboreus происходит в течение всего периода диапаузы за счет липидных запасов.
5. В комплексе оборонительных реакций планктонных копепод важнейшими оказываются тактики, позволяющие предотвратить встречу с хищником. Сезонные вертикальные миграции и диапауза, как наиболее радикальный способ ухода копепод от хищников, в значительной степени определяют тип жизненного цикла. Миграции сильнее выражены среди наиболее уязвимых для хищников и репродуктивно ценных для популяции особей.
6. Копеподы с жизненным циклом, включающим наиболее длительную диапаузу (Calanus hyperboreus, Calanoides acutus), способны накапливать необходимый запас липидов (более 50% сухой массы тела) в короткий период «цветения» фитопланктона благодаря их необычайно высокой удельной скорости потребления пищи. Энергетическая ценность их резервов обусловлена также высокой (до 90%) долей в них восковых эфиров со значительным содержанием полиненасыщенных жирных кислот.
7. Различия в жизненных циклах антарктических копепод (Calanoides acutus, Calanus propinquus, Rhincalanus gigas), связанные с диапаузой, выявлены на основании впервые полученных одновременных оценок скорости метаболизма в «поверхностной» и «глубинной» субпопуляциях в переходный период от осени к зиме. C. acutus и C. propinquus в глубинных слоях находились в диапаузе, так как их метаболизм был в 4-8 раз снижен по сравнению с особями из поверхностного слоя. У погрузившихся в глубину R. gigas метаболизм оставался на том же уровне, что и в «поверхностной» субпопуляции, т. е., они были в активном состоянии.
8. Стратегия жизненного цикла копепод не является видоспецифичной. Один и тот же вид может использовать одну или несколько стратегий. Например, впадать в диапаузу или нет (Calanus propinquus), размножаться до начала питания или одновременно с питанием (C. glacialis).
9. Стратегия жизненного цикла связана с размером копепод. У крупных копепод больше возможностей выбора между стратегиями, чем у мелких. Их спектр питания обычно шире, чем у мелких копепод, а удельные энергетические затраты на двигательную активность ниже. Они могут уходить в диапаузу в зависимости от уровня жировых резервов или оставаться активными при благоприятных локальных условиях среды (много корма и мало хищников).
Основные публикации по теме диссертации
1. Пастернак A. Ф. 1974. Усвояемость пищи и коэффициент использования энергии усвоенной пищи на рост япономорской мизиды Neomysis mirabilis (Czernjavsky). Вестник Московского Университета, сер. Биология, почвоведение, 1: 15-17
2. , , 1974. Совмещение радиоуглеродного метода с математическим моделированием для оценки продуктивности морского зоопланктона. Океанология, 14(2): 227-235
3. 1976. Некоторые данные о росте и размножении Tisbe furcata (Copepoda, Harpacticoida) в Черном море. Зоологический журнал,:
4. Пастернак A. Ф. 1977. Изменение некоторых показателей трансформации энергии в процессе роста мизиды Neomysis mirabilis. Океанология, 17(4): 713-718
5. Пастернак A. Ф., 1980. Скорость дыхания мелкого зоопланктона в районе Перуанского апвеллинга. Стр. 190-198 в кн.: (ред.) «Пелагические экосистемы Перуанского района». Наука, Москва
6. Pasternak A. F., Averianov A. A. 1980. Respiration of minute forms of zooplankton and net growth efficiency of some copepods in the Peruvian upwelling region. Pol. Arch. Hydrobiol., 27(4): 485-496
7. Пастернак A. Ф., , 1981. Потребление гетеротрофных жгутиковых планктонными организмами. Океанология, 21(2): 375-379
8. Пастернак A. Ф. 1982. Сезонные изменения численности и биомассы зоопланктона у побережья северного Кавказа. Стр. 78-95 в кн.: (ред.) «Сезонные изменения в планктоне Черного моря». Наука, Москва
9. , , 1982. Продукционные и деструкционные характеристики планктонного сообщества северо-восточной части Черного моря в 1978. Стр. 139-177 в кн.: (ред.) «Сезонные изменения в планктоне Черного моря». Наука, Москва
10. , , 1983. Изменение скорости дыхания массовых видов зоопланктона Черного моря в течение года. Океанология, 23(1): 145-149
11. 1984. Питание веслоногих рачков рода Oncaea (Copepoda: Cyclopoida) в юго-восточной части Тихого океана. Океанология, 24(5): 808-812
12. , , 1984. Роль паразитических водорослей рода Blastodinium в экологии планктонных копепод. Океанология, 24(6): 994-998
13. , 1984. Размерный состав пищи представителей различных трофических групп зоопланктона восточной части Тихого океана. Стр. 247-256 в кн. «Фронтальные зоны юго-восточной части Тихого океана (ред. , ) Наука, Москва
14. , , 1987. Использование лазерного флуориметра для определения содержания растительных пигментов в кишечниках планктонных животных. Океанология, 27(5): 852-856
15. , , 1987. Определение выедания фитопланктона зоопланктоном в Балтийском море весной 1984. Стр. 343-369 в кн.: -Мишке (ред.) «Экосистемы Балтийского моря в мае-июне 1984 г. Материалы 39-го рейса нис «Академик Курчатов». ИОАН, Москва
16. , 1988. Исследование деградации фекальных пеллет при симуляции их погружения в толще воды. Океанология, 28(2): 316-321
17. 1988. Состав пищи у доминирующих планктонных животных. Стр. 251-254 в кн. : , (ред.) «Экосистемы субантарктической зоны Тихого океана». Наука, Москва
18. Pasternak A. F., Drits A. V. 1988. Possible degradation of chlorophyll-derived pigments during gut passage of herbivorous copepods. Mar. Ecol. Progr. Ser., 49: 187-190
19. , 1989. Влияние концентрации пищи на активность питания и время прохождения пищи через кишечник у растительноядных копепод в зонах апвеллингов. Океанология, 29(2): 309-315
20. , , 1990. Особенности физиологии некоторых интерзональных видов копепод. Журнал Общей Биологии, 51(3): 382-392
21. , , 1990. Исследование суточного ритма питания антарктической копеподы Calanus propinquus. Океанология, 30(6):
22. , , 1990. Питание копепод-фитофагов в районах апвеллингов. Суточные ритмы, рационы и выедание фитопланктона. Океанология, 30(2): 311-319
23. , , 1991. Зависит ли время прохождения пищи через кишечник копепод от концентрации пищи в среде? Океанология, 31(1): 47-54
24. , 1991. О возможности разрушения растительных пигментов (хлорофилла и его производных) при прохождении пищи через кишечник растительноядных копепод. Стр. 306-313 в кн.: (ред.) «Исследования фитопланктона в рамках мониторинга Балтийского и других морей СССР». Гидрометеоиздат, Москва
25. Flint M. V., Drits A. V., Pasternak A. F. 1991. Characteristic features of body composition and metabolism in some interzonal copepods. Mar. Biol., 111: 199-205
26. Drits A. V., Pasternak A. F., Kosobokova K. N. 1993. Feeding, metabolism and body composition of the Antarctic copepod Calanus proninquus Brady with special reference to its life cycle. Polar Biol., 13(1): 13-21
27. , 1993. Питание массовых видов антарктического зоопланктона в летний период. Стр. 250-259 в кн. «Пелагические экосистемы Южного океана» (ред. ). Наука, Москва
28. Drits A. V., Pasternak A. F., Kosobokova K. N. 1994. Physiological characteristics of the Antarctic copepod Calanoides acutus during late summer in the Weddell Sea. Hydrobiologia, 292/293: 201-207
29. Pasternak A. F. Gut fluorescence in herbivorous copepods: an attempt to justify the method. 1994. Hydrobiologia, 292/293: 241-248
30. Pasternak A. F., Kosobokova K. N., Drits A. V. 1994. Feeding, metabolism and body composition of the dominant Antarctic copepods with comments on their life *****ssian Journ. Aquatic Ecology, 3(1): 49-62
31. Pasternak A. F., Huntingford F. A., Crompton D. W.T. 1995. Changes in metabolism and behaviour of the freshwater copepod Cyclops strenuus abyssorum infected with Diphyllobothrium spp. Parasitology, 110: 395-399
32. Pasternak A. F. 1995. Gut contents and diel feeding rhythm in dominant copepods in the ice-covered Weddell Sea, March 1992. Polar Biology, 15: 583-586
33. Pasternak A. F., Arashkevich E. G., Semenova T. N. 1996. Summer delay in the development of Eudiaptomus graciloides: is this a diapause? Russian Journal of Aquatic Ecology, 5(1-2): 39-47
34. , , 1998. Связь вертикального распределения и стратегии жизненного цикла Eudiaptomus graciloides (Lilljeborg 1888) в озере Глубоком. Биология внутренних вод, 1: 29-38.
35. Pasternak A. F. and Arashkevich E. G. 1999. Resting stages in the life cycle of Eudiaptomus graciloides (Lill) (Copepoda:Calanoida) in Lake Glubokoe. Journal of Plankton Research, 21: 309-325
36. Pasternak A. F., Pulkkinen K., Mikheev V. N., Hasu T. and Valtonen E. T. 1999. Factors affecting abundance of Triaenophorus infection in Cyclops strenuus, and parasite-induced changes in host fitness. Int. Journ for Parasitology 29: .
37. Reigstad M., Wassmann P., Ratkova T., Arashkevich E., Pasternak A. and Oeygarden S. parison of the springtime vertical export of biogenic matter in three northern Norwegian fjords. Mar. Ecol. Progr. Ser. 201: 73-89.
38. Pasternak A. F., Mikheev V. N. and Valtonen E. T. 2000. Life history characteristics of Argulus foliaceus L. (Crustacea: Branchiura) populations in Central Finland. Ann. Zool. Fennici 37: 25-35.
39. Pulkkinen K., Pasternak A. F., Hasu T. and Valtonen E. T. 2000. Effect of Triaenophorus crassus (Cestoda) infection on behavior and susceptibility to predation of the first intermediate host Cyclops strenuus. J. Parasitology 86(4): 664-670.
40. Pasternak A., Arashkevich E., Wexels Riser C., Rat’kova T. and Wassmann P. 2000. Seasonal variation in zooplankton and suspended faecal pellets in the subarctic fjord Balsfjorden, northern Norway, in 1996. Sarsia 85: 439-452.
41. Pasternak A.., Arashkevich E., Tande K., Falkenhaug T. 2001. Seasonal changes in feeding, gonad development and lipid stores in Calanus finmarchicus and C. hyperboreus from Malangen, northern Norway. Marine Biology 138: .
42. Pasternak A., Schnack-Schiel S. B. 2001. Seasonal feeding patterns of the dominant Antarctic copepods Calanus propinquus and Calanoides acutus in the Weddell Sea. Polar Biology: 771-784
43. Mikheev V. N., Pasternak A. F., Valtonen E. T., Lankinen Y. 2001. Spatial distribution and hatching of overwintered eggs of a fish ectoparasite, Argulus coregoni Thorell (Crustacea: Branchiura). Diseases of Aquatic Organisms: 123-128
44. Pasternak A., Schnack-Schiel S. B. 2001. Feeding patterns of dominant Antarctic copepods: an interplay of diapause, selectivity and availability of food. Hydrobiologia 453/454: 25-36
45. Pasternak A., Wexels Riser C., Arashkevich E., Rat’kova T., Wassmann P. 2002. Calanus spp. grazing affects egg production and vertical carbon flux (the marginal ice zone and open Barents Sea). Journal of Marine Systems V.38: 147-164
46. Arashkevich E., Wassmann P., Pasternak A., Wexels Riser C. 2002. Seasonal and spatial changes in biomass, structure and development progress of the zooplankton community in the Barents Sea. Journal of Marine Systems V. 38: 125-145
47. , 2002. Пищевая реакция мальков окуня Perca fluviatilis и плотвы Rutilus rutilus на присутствие хищника. Доклады АН, Общая Биология, т. : 842-844
48. Wexels Riser C., Wassmann P., Olli K., Pasternak A., Arashkevich E., 2002. Seasonal variation in production, retention and export of zooplankton faecal pellets in the marginal ice zone and central Barents Sea. Journal of Marine Systems V. 38: 175-188
49. Olli K., Wexels Riser C., Wassmann P., Ratkova T., Pasternak A., Arashkevich E., Pasternak A. 2002. Seasonal variation in vertical flux of biogenic matter in the marginal ice zone and the central Barents Sea. Journal of Marine Systems V. 38: 189-204
50. Mikheev V. N., Pasternak A. F., Valtonen E. T.. 2003. How do fish ectoparasites Argulus spp. (Crustacea: Branchiura) match with their hosts at the behavioural and ecological scales? Журнал Общей Биологии, 64(3): 238-47
51. Hakalahti T., Pasternak A. F., Valtonen E. T. 2004. Seasonal dynamics of egg laying and egg-laying strategy of the ectoparasite Argulus coregoni (Crustacea: Branchiura). Parasitology, 128: 655-660.
52. Pasternak A., Tande K. S., Arashkevich E., Melle W. 2004. Reproductive patterns of Calanus finmarchicus at the Norwegian midshelf in 1997. Journ. of Plankton Res.,: 839-849.
53. Pasternak A., Mikheev V., Valtonen E. T. 2004. Growth and development of Argulus coregoni (Crustacea: Branchiura) on salmonid and cyprinid hosts. Diseases of Aquatic Organisms, 58: 203-207.
54. , , 2004. Адаптивное значение размерно-полового диморфизма у эктопаразита рыб Argulus coregoni (Crustacea: Branchiura). Доклады АН, Общая Биология, т. : 1-4.
55. Arashkevich E. G., Tande K. S., Pasternak A. F., Ellertsen B. 2004. Seasonal moulting patterns and the generation cycle of Calanus finmarchicus in the NE Norwegian Sea, as inferred from gnathobase structures, and the size of gonads and oil sacs. Marine Biology, 146: 119-132.
56. Pasternak A., Wassmann P., Wexels Riser C. 2005. Does mesozooplankton respond to episodic phosphorus inputs in the eastern Mediterranean? Deep-Sea Research II: Topical studies in oceanography, 52:
57. Mikheev V. N., Wanzenböck J., Pasternak A. F. 2006. Effects of predator-induced visual and olfactory cues on 0+ perch (Perca fluviatilis L.) foraging behaviour. Ecology of Freshwater Fish, 15: 111-117.
58. Pasternak A. F., Mikheev V. N., Wanzenböck J. 2006. How plankton copepods avoid fish predation: from individual responses to variations of the life cycle. Вопросы ихтиологии (Journal of Ichthyology), 46 (Suppl. 2): 220-226.
59. Mikheev V. N., Pasternak A. F.2006. Defense behavior of fish against predators and parasites. Вопросы ихтиологии (Journal of Ichthyology), 46 (Suppl. 2): 173-179.
60. Olli K., Wassmann P., Reigstad M., Rat’kova T., Arashkevich E., Pasternak A., Matrai P., Knulst J., Tranvik R., Klais R., Jacobsen A. 2007. The fate of production in the central Arctic Ocean – top-down regulation by zooplankton expatriates? Progress in Oceanography 72(1): 84-113.
61. Pasternak, A. F. and Schnack-Schiel, S. B. 2007. Feeding of Ctenocalanus citer in the eastern Weddell Sea: low in summer and spring, high in autumn and winter. Polar Biology, 30: 493-501.
На заключительном этапе работа выполнялась при финансовой поддержке Российского фонда фундаментальных исследований (проект № )

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
