

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
РЫБКО Вера Александровна
ИССЛЕДОВАНИЕ МОЛЕКУЛЯРНЫХ МЕХАНИЗМОВ
RAL-ОПОСРЕДОВАННОЙ СТИМУЛЯЦИИ МЕТАСТАЗИРОВАНИЯ
Специальность 14.00.14 – онкология
АВТОРЕФЕРАТ
Диссертации на соискание ученой степени
кандидата биологических наук
Москва
2009
Работа выполнена в Лаборатории регуляции клеточных и вирусных онкогенов НИИ Канцерогенеза Учреждения Российской академии медицинских наук Российский Онкологический Научный Центр имени РАМН
Научный руководитель:
Кандидат биологических наук
Официальные оппоненты:
Доктор биологических наук
Доктор биологических наук
Ведущая организация: Научно-исследовательский институт физико-химической биологии им. ,
МГУ им.
Защита диссертации состоится «___» _______ 2009 года в ___ часов на заседании диссертационного совета (Д.001.017.01) РОНЦ имени РАМН Москва, Каширское шоссе, 24.
С текстом диссертации можно ознакомиться
в библиотеке РОНЦ имени РАМН
Автореферат разослан «___» _________ 2009 года.
Ученый секретарь
диссертационного совета
д. м.н., профессор
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы
Канцерогенез представляет огромный интерес для фундаментальной молекулярной биологии, поскольку причиной возникновения всех опухолевых заболеваний являются нарушения в сложных механизмах регуляции клеточной жизнедеятельности, до сих пор до конца не изученных даже в норме. Одной из актуальнейших проблем является идентификация белков, активность которых изменяется при прогрессии заболевания. Исследование таких нарушений может в будущем привести к разработке новых мишеней для направленной терапии.
В настоящее время не вызывает сомнения огромная роль онкогена Ras как в инициации, так и в прогрессии злокачественных опухолей человека. Активированный Ras может передавать сигнал на несколько сигнальных путей, участие каждого из которых в трансформации клеток и опухолевой прогрессии может быть различным. Среди таких путей наиболее известны Ras-Raf-ERK, Ras-PI3K-PDK1 и Ras-RalGEFs-Ral сигнальные каскады. До недавнего времени важнейшая роль в онкогенезе приписывалась Raf-ERK пути, однако современные исследования показали, что он играет столь большое значение в первую очередь в моделях мышиных клеток, в то время как для трансформации человеческих клеток более важным может являться путь активации малых ГТФаз Ral. Таким образом, изучение механизмов действия двух высокогомологичных белков RalA и RalB является крайне актуальным для понимания механизмов онкогенеза у человека.
Одним из широко-используемых современных подходов к исследованию вклада отдельных сигнальных каскадов, ассоциированных с малыми ГТФазами (или G-белками) является использование эффекторных мутантов, способных избирательно активировать определенные белки. Мутации в эффекторном домене малых ГТФаз, ответственном за взаимодействие с белками-партнерами, могут прекращать активацию того или иного эффектора. При суперэкспрессии таких генов (далее – эффекторных мутантов), прекращается передача сигнала, опосредованного взаимодействием малого G-белка с соответствующей мишенью, что позволяет исследователям оценивать вклад белков-партнеров в исследуемые характеристики клеток.
В лаборатории регуляции вирусных и клеточных онкогенов ранее показано, что онкогенная форма Ha-Ras способна усиливать спонтанное органоспецифическое метастазирование в легкие трансформированных v-src эмбриональных фибробластов Сирийского хомяка (клеточной линии Het-SR). Более того, при исследовании вклада эффекторных мутантов Ha-Ras, способных активировать в отдельности PI3K-, Raf - или RalGDS-ассоциированные сигнальные каскады (V12C40, V12S35 и V12G37 соответственно), показано, что именно активация последнего является основным событием, приводящим к усилению метастазирования в данной модели (Tchevkina et al., 2005).
Белки RalA и RalB высокогомологичны друг другу как по последовательности, так и по спектру белков-партнеров, основными из которых являются RalBP1, PLD и комплекс экзоциста. Активация этих мишеней может регулировать такие ключевые внутриклеточные процессы, как мембранный транспорт, передача сигнала, регуляция экспрессии генов, прохождение клеточного цикла и др. До настоящего времени не проводилось комплексной оценки вклада взаимодействия белков Ral с отдельными эффекторами в такие изменения. Таким образом, данное исследование вклада RalA и RalB в различные характеристики злокачественных клеток, а также молекулярных механизмов, с помощью которых белки Ral оказывают влияние на различные этапы опухолевой прогрессии, и, в конечном счете, на метастазирование, крайне актуально и представляют значительный научный интерес. Кроме того, вопрос о том, активация каких Ral-опосредованных сигнальных каскадов может приводить к изменению свойств злокачественных клеток остается открытым.
Цель и задачи исследования
Цель данной работы - изучить влияние белков RalA и RalB на потенциально ассоциированные с метастазированием характеристики трансформированных клеток.
Для достижения цели были поставлены следующие экспериментальные задачи:
1. Получить клеточные линии Het-SR, стабильно экспрессирующие ГТФ-связанные формы RalA и RalB, а также их эффекторные мутанты, неспособные активировать каждый из трех наиболее хорошо изученных Ral-ассоциированных сигнальных каскадов.
2. Изучить характеристики полученных линий in vitro:
- динамику пролиферации
- подвижность
- способность к колониеобразованию в условиях разреженной популяции и неприкрепленного роста
- способность к инвазии и движению по градиенту факторов роста.
3. Исследовать влияние различных форм Ral на секрецию протеаз, ответственных за ремоделирование внеклеточного матрикса, а также на внутриклеточную передачу сигнала.
4. Исследовать влияние различных форм белков Ral на туморогенность и спонтанную метастатическую активность клеток Het-SR.
5. Сопоставить результаты исследований влияния различных форм белков Ral на клетки in vivo и in vitro.
Научная новизна и практическая значимость исследования
Модель клеточных линий, полученных в данной работе, позволяет оценивать вклад двух близкородственных ГТФаз RalA и RalB, а также их основных белков партнеров в изменении характеристик трасформированных клеток не только in vitro, но и in vivo, начиная с формирования первичного очага опухолевого роста до развития метастазов в отдаленных органах. Данная модель состоит из производных v-src-трансформированных эмбриональных фибробластов сирийского хомяка. Родительская линия Het-SR является высокотуморогенной на сингенных животных, однако обладает слабым метастатическим потенциалом при подкожном введении.
Полученные в работе результаты расширяют представления о роли малых ГТФаз Ral в канцерогенезе и о Ral-опосредованных сигнальных каскадах, активация которых необходима для приобретения клетками злокачественных характеристик. Данное исследование свидетельствуют в пользу активного участия белков Ral и Ral-ассоциированных сигнальных каскадов в различных этапах прогрессии опухолей, и в том числе в метастазировании.
Работа носит в первую очередь теоретический характер, однако полученные данные о роли данных белков в опухолевой прогрессии могут быть использованы при дальнейшем изучении участия белков Ral как на модельных системах, так и в клинических исследованиях. Возможно, что такие исследования приведут к открытию новых диагностических маркеров и мишеней для таргетной терапии опухолей.
Положения, выносимые на защиту
1. Малые ГТФазы RalA и RalB (в большей степени) стимулируют спонтанную метастатическую активность клеток. Эффекторный мутант RalA, неспособный взаимодействовать с комплексом экзоциста является наиболее активным по данному параметру. Взаимодействие RalB с PLD является критичным для стимуляции метастазирования.
2. Активные формы белков Ral, а также их эффеторные мутанты могут в той или иной степени влиять на деление клеток, как при стандартных методах культивирования, так и в условиях неприкрепленного роста или разреженной популяции. Они оказывают влияние на подвижность клеток, а также могут стимулировать миграцию по градиенту сыворотки и инвазию через мембраны, покрытые матригелем.
3. Белки Ral активно участвуют во внутриклеточной передаче сигнала, стимулируя как активацию различных сигнальных каскадов, так и экспрессию белков, потенциально ассоциированных с опухолевой прогрессией. В частности, активная форма RalA стимулирует секрецию протеазы, влияющей на ремоделирование внеклеточного матрикса uPA и экспрессию поверхностного гликопротеина CD24, а RalB снижает экспрессию белка кавеолин-1.
Личное участие автора в получении результатов,
изложенных в диссертации
Все исследования проведены лично автором. В ходе исследования получены клеточные линии, стабильно экспрессирующие различные формы белков Ral. Проведен анализ спонтанной метастатической активности и туморогенности всей панели лний на экспериментальных животных, а также изучены клеточные характеристики in vitro.
Апробация работы
Диссертация апробирована и рекомендована к защите 24 сентября 2008 года на совместной научной конференции лабораторий регуляции клеточных и вирусных онкогенов, молекулярной биологии вирусов, вирусного канцерогенеза, цитогенетики, механизмов прогрессии эпителиальных опухолей и противоопухолевого иммунитета НИИ Канцерогенеза ГУ РОНЦ им. РАМН. Материалы работы докладывались на конференциях «Second International Congress on Molecular Staging of Cancer» в 2005 г. (Гейдельберг, Германия), «Методы культивирования клеток» в 2009 г. (Санкт-Петербург).
Публикации
Результаты диссертационной работы отражены в 8-ми печатных работах соискателя, в том числе 2 статьи и 3 тезиса опубликованы в журналах, рекомендованных ВАК МОН РФ соискателям ученой степени кандидата биологических наук.
Структура и объём диссертации
Диссертация изложена на 126 страницах машинописного текста, содержит 44 рисунка, 3 таблицы. Состоит из глав: «Введение», «Обзор литературы», «Материалы и методы», «Результаты», «Обсуждение», «Выводы», «Список использованной литературы». Список литературы содержит 162 источника, в том числе 2 в отечественных рецензируемых изданиях.
СОДЕРЖАНИЕ РАБОТЫ
Материалы и методы
В работе использованы следующие методы исследования: молекулярное клонирование, полимеразная цепная реакция (ПЦР), трансфекция эукариотических клеток, инфицирование клеток ретровирусными векторами, анализ белков с помощью вестерн-блот гибридизации и зимографий, анализ динамики роста клеток с использованием МТТ, анализ «зарастания раны» клеточными линиями in vitro (wound healing assay), тест на образование колоний в условиях разреженной популяции, тест на образование колоний в полужидкой среде, тест на миграцию клеток по градиенту концентраций факторов роста, тест на инвазию in vitro, определение спонтанной метастатической активности (СМА) и туморогенности клеток на лабораторных животных, а также статистическая обработка данных.
Результаты исследования и обсуждение
1. Клеточные линии, использованные в работе
Для изучения влияния белков Ral на характеристики трансформированных клеток получена панель стабильных клеточных линий-производных Het-SR с помощью ретровирусных векторов (см. рисунок 1). Для проведения данного исследования использовались гены, несущие следующие мутации:
- RalA/B V23 – мутантная форма с аминокислотной заменой в 23-м положении (глицин на валин), конститутивно связанная с ГТФ и, таким образом, активирующая все Ral-ассоциированные сигнальные каскады;
- RalA/B V23 deltaN11 – активная форма с делетированным аминоконцевым участком, ответственным за связывание с PLD;
- RalA/B V23 D49N – активная форма с аминокислотной заменой в участке эффекторной петли в 49-м положении (аспарагиновая кислота на аспарагин), неспособная связываться с RalBP1;
- RalA/B V23 D49E – активная форма с аминокислотной заменой в 49-м положении (аспарагиновая кислота на глутаминовую кислоту), неспособная связываться с белком Sec5, входящим в состав комплекса экзоциста.
Таким образом, в данной работе оказалось возможным оценить вклад отсутствия активации каждого из трех основных эффекторов как RalA, так и RalB в те или иные молекулярно-биологические характеристики клеточной линии Het-SR.
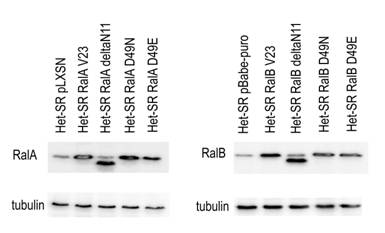
Рисунок 1. Вестерн-блот анализ экспрессии RalA и RalB в полученных клеточных линиях. Антитела к тубулину использовались в качестве контроля
2. Характеристики полученных клеточных линий in vivo
Полученные клеточные линии были исследованы на туморогенность и спонтанную метастатическую активность на лабораторных животных. Для этого они были введены подкожно (максимально по 20000 клеток) и через 2 месяца оценивался размер опухолей, минимальная прививочная доза, а также проводилось гистологическое исследование количества метастазов в легких (см. рисунок 2)
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Рисунок 2. Количество гистологически-верифицированных метастазов в легких животных. Звездочка – статистически-достоверное отличие от контроля
Статистическая обработка данных показала, что RalA стимулирует спонтанную метастатическую активность клеток (1.910 ± 0.7663 vs 16.25 ± 4.358, p<0,05). Эффекторные мутанты RalA с заблокированными взаимодействиями с RalBP1 или с PLD1 также статистически значимо повысили метастазирование клеточной линии Het-SR. Однако эффекторный мутант RalA D49E, отменяющий ассоциацию RalA с комплексом экзоциста, повысил метастазирование более чем в 4 раза по сравнению с полностью активной формой белка.
Активная форма RalB значительно стимулировала образование легочных метастазов по сравнению с контрольной линией Het-SR pBabe-puro (70.79 ± 20.65 vs 5.300 ± 1.499, p<0.05). При этом эффекторный мутант RalB, неспособный взаимодействовать с PLD статистически значимо не повлиял на метастатический потенциал опухолей (12.00 ± 5.379). Эффекторные мутанты RalB D49N и D49E также стимулировали метастазирование (45.65 ± 14.75 и 35.65 ± 12.05 соответственно). Достоверных различий между полностью активной формой и эффекторными мутантами с аминокислотными заменами в 49-м положении выявлено не было. Однако имеется тенденция к снижению количества метастазов при ингибировании взаимодействия RalB c RalBP1 или с комплексом экзоциста.
Таким образом, RalA и RalB по-разному влияют на метастатический потенциал клеток. Для стимуляции образования очагов вторичного роста необходимо взаимодействие RalB с PLD, а RalA не должен взаимодействовать с комплексом экзоциста. Возможно, что конститутивная связь RalA с ГТФ тормозит определенные процессы в клетке (в частности сборку экзоциста), а эффекторный мутант, неспособный связываться с Seс5 дает возможность этому мультибелковому комплексу влиять на метастатический потенциал клеток.
3. Характеристики полученных клеточных линий in vitro
Одной из основных характеристик трансформированных клеток является повышенная скорость деления. По литературным данным известно, что белки Ral могут влиять на пролиферацию клеток, в частности увеличивая экспрессию CyclinD1 или подавляя активность ингибитора клеточного цикла p27kip. При исследовании влияния активных форм RalA и RalB на пролиферацию клеток линии Het-SR оказалось, что они в значительной степени ускоряют скорость деления (см. рисунок 3). Более того, анализ данных по динамике пролиферации клеток, экспрессировавших эффекторные мутанты белков Ral, показал, что взаимодействие RalA с комплексом экзоциста, а также с белком RalBP1 является необходимым условием для RalA-зависимой стимуляции деления клеток (эффекторные мутанты RalA D49E и RalA D49N не оказывали влияния на скорость деления). Взаимодействие RalA с PLD, скорее всего, вносит определенный вклад в ускорение деления клеток, однако он не столь значителен. Необходимо отметить, что в современной научной литературе есть данные, указывающие на возможное влияние активности RalBP1 на пролиферацию.
![]()
![]()
![]()
![]()
![]()
![]()
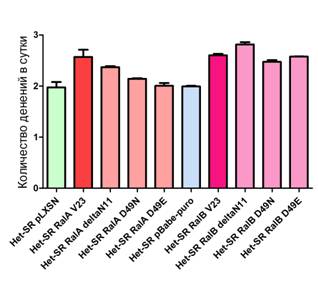
Рисунок 3.Среднее количество удвоений в сутки исследованных клеточных линий. Звездочка – статистически достоверное отличие от контроля (p<0.05)
Данные о скорости пролиферации клеток, экспрессирующих различные формы RalB, указывают на то, что эта характеристика не зависит от взаимодействия с каким-то одним из основных эффекторов Ral (все эффекторные мутанты RalB приводят к ускоренной пролиферации клеток по сравнению с контролем). Этот факт может объясняться двумя гипотезами:
- для RalB-опосредованной ускоренной пролиферации достаточно активации 2-х эффекторов;
- активация не учтенного в нашей работе белка-партнера, взаимодействие с которым сохраняется во всех использованных вариантах эффекторных мутантов.
Тест на формирование колоний при отсутствии микроокружения (тест на колониеобразование) отражает способность клеток пролиферировать без «поддержки» соседних клеток (см. Рисунок 4). Оказалось, что практически все исследованные линии обладают этой способностью, что говорит о большой степени их автономности от микроокружения. Только клеточная линия Het-SR RalA D49E, в которой нарушено взаимодействие RalA с комплексом экзоциста, не отличается по этой характеристике от контроля. Учитывая то, что нами не выявлено отличий этой линии по скорости пролиферации в условиях стандартного культивирования от линии Het-SR pLXSN, можно сделать вывод, что взаимодействие RalA c экзоцистом играет важную роль в пролиферации клеток как в условиях бедного микроокружения, так и при стандартной культивации.
С другой стороны, клеточная линия Het-SR RalA D49N также не отличалась по скорости пролиферации от Het-SR pLXSN в стандартных условиях, в то время как в тесте на образование колоний она формировала большие по площади фокусы роста. Таким образом, можно предположить, что взаимодействие RalA с RalBP1 не влияет на рост клеток в таком тесте, но необходимо для ускоренного роста в стандартных условиях. Активные формы RalB, также как и эффекторные мутанты этой ГТФазы, стимулировали колониеобразование в данном эксперименте.
![]()
![]()
![]()
![]()
![]()
![]()
![]()

Рисунок 4. Средний размер отдельных колоний полученных клеточных линий. Звездочка – статистически достоверное отличие от контроля (p<0.05)
В ходе работы был поставлен вопрос, опосредована ли Ral-зависимая стимуляция пролиферации усилением экспрессии CyclinD1. Как видно из рисунка 5 активные формы Ral действительно усиливают экспрессию этого гена. Однако в нашей модели RalB-опосредованная активация RalBP1 скорее всего играет важную роль в усилении экспрессии CyclinD1, так как лишь эффекторный мутант с нарушенным взаимодействием между этими белками (RalB D49N) не отличался от контроля. Нами не было выявлено достоверных отличий в количестве CyclinD1 в клеточных линиях с экзогенной экспрессией эффекторных мутантов RalA по сравнению с контролем.
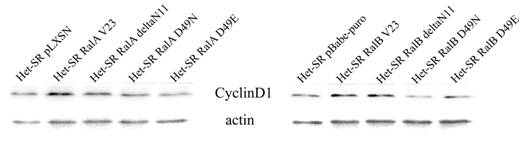
Рисунок 5. Вестерн-блот анализ экспрессии CyclinD1 (в качестве контроля использовались антитела к β-актину)
При исследовании подвижности клеток с заблокированным митозом (с помощью Митомицина С) в тесте на «зарастание раны in vitro» оказалось, что RalB лишь немного ускоряет движение клеток, тогда как клетки линии Het-SR RalA V23 мигрируют в «рану» в 3 раза быстрее, чем контрольные. Более того, интересно, что эффекторные мутанты RalA D49N (эта форма лишь немного ускоряла движение клеток) и RalA D49E (совершенно неотличима от контроля) не влияли на подвижность клеток, что указывает на значимость взаимодействия этой малой ГТФазы c RalBP1 и, в особенности, с экзоцистом в этом процессе (см. рисунок 6). Таким образом, представляется, что RalA играет существенную роль в стимуляции миграционной активности клеток в данной модельной системе, и что эта функция малой ГТФазы осуществляется, скорее всего, через взаимодействие с комплексом экзоциста.
![]()
![]()
![]()
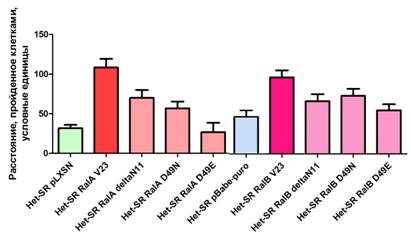
Рисунок 6. Миграция клеток, экспрессирующих различные формы белков Ral, в среде c 10% сыворотки и 1.5 мкг/мл Звездочка – статистически достоверное отличие от контроля (p<0.05)
Направление миграции опухолевых (как и нормальных) клеток в организме определяется градиентом различных факторов, которые задают направление движения. В упрощенной модели этого процесса in vitro рассматривается миграция клеток через поры мембраны по градиенту сыворотки в камерах Бойдена. Результаты этого теста могут отображать способность клеток к направленному движению. Исследование такого движения клеток Het-SR, экспрессирующих различные формы белков Ral, показало, что RalB, в целом, в большей степени, чем RalA стимулирует направленное движение, и что каждый из шести исследованных эффекторных мутантов был менее эффективен, чем конститутивно-активные формы белков Ral (см. рисунок 7). При этом основным партнером исследуемых ГТФаз в стимуляции миграции по градиенту сыворотки является комплекс экзоциста. Эти результаты в некоторой степени соотносятся как с исследованием миграционной способности в тесте на зарастание раны, так и с литературными данными.
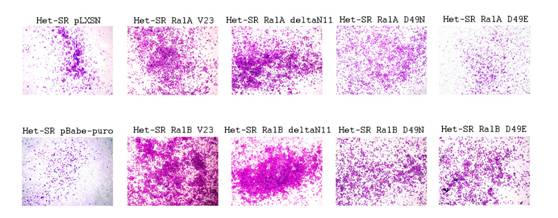
Рисунок 7. Миграция клеток по градиенту сыворотки в камере Бойдена.
Способность к инвазии – ключевая характеристика злокачественных клеток. Она заключается в преодолении барьера капсулы, окружающей опухоль, а также проникновении опухолевых клеток в лимфатические и кровеносные сосуды. In vitro способность к инвазии оценивают с помощью так-называемых камер Бойдена, дно которых покрыто матригелем. В них моделируется ситуация, при которой трансформированные клетки оказываются в состоянии голодания по факторам роста, и для того, чтобы их получить, клеткам необходимо преодолеть барьер, состоящий из аналога ВКМ (внеклеточного матрикса) – матригеля. Такой тест отражает способность клеток к инвазии в целом, а также характеризует направленное движение и разрушение внеклеточного матрикса.
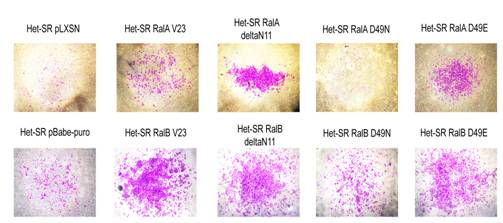
Рисунок 8. Фотографии внешней стороны мембраны после проведения теста на инвазию in vitro
При исследовании способности к инвазии in vitro оказалось, что активная форма RalA лишь немного усиливает инвазивные способности клеток Het-SR, в то время как RalB значительно повышает количество клеток, преодолевших барьер (см рисунок 8). Тем не менее, эффекторные мутанты RalA, неспособные взаимодействовать с PLD или с комплексом экзоциста, усиливают инвазивный потенциал клеток Het-SR. Таким образом, можно предположить, что отсутствие взаимодействия RalA с этими партнерами является ключевым в этой клеточной характеристике. Более того, эффекторные мутанты как RalA, так и RalB, неспособные взаимодействовать с RalBP1, практически не отличались по этой характеристике от контрольных линий, в то время как все остальные клеточные линии, экспрессирующие RalB, обладают значительно более высоким инвазивным потенциалом.
4. Влияние белков Ral на активацию сигнальных каскадов и экспрессию белков, ассоциированных с метастазированием
Опухолевая строма в целом, и внеклеточный матрикс в частности, оказывает огромное влияние на прогрессию злокачественных новообразований. Взаимодействие трансформированных клеток с микроокружением может стимулировать или подавлять многие процессы, ассоциированные с инвазией и метастазированием. Злокачественные клетки активно ремоделируют свое микроокружение, в том числе разрушая ВКМ при помощи протеолитических ферментов. При активации систем деградации внеклеточного матрикса опухолевые клетки могут приобрести способность инвазировать в окружающую ткань, при этом также могут высвобождаться факторы роста, «заякоренные» в нем.
Исследование активности одной из таких секретируемых протеаз, Урогиназа-подобный активатор плазминогена (uPA), в кондиционированных средах от полученных клеточных линий выявило интересную закономерность. Активная форма RalA приводит к некоторому увеличению уровня секреции этой протеазы (см. рисунок 9). В клеточных линиях, экспрессировавших эффекторные мутанты RalA D49N и D49E, как и в RalB-трансфектантах, секреция uPA не повышалась, что говорит о ключевой роли взаимодействия RalA с RalBP1 и с комплексом экзоциста в стимуляции активности этой протеазы.
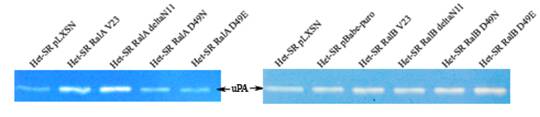
Рисунок 9. Анализ активности uPA в кондиционированных средах от полученных линий
Показано, что MMP (матриксные металлопротеазы) влияют на прогрессию опухолевых заболеваний в целом, и на метастазирование в частности. Исследования уровня секреции матриксных металлопротеаз, обладающих желатиназной активностью, показало, что в кондиционированных средах от клеточных линий-производных Het-SR наиболее активной протеазой является MMP2 (см. рисунок 10). При этом было выявлено слабое усиление активности ММP9 и MMP1 при экспрессии активной формы RalA, неспособной взаимодействовать с PLD. В то же время эффекторный мутант RalB D49E значимо изменил внеклеточную активность этих протеаз, что говорит о сложности и неоднозначности Ral-зависимой регуляции секреции MMP и о различиях в необходимых партнерах для этого процесса.
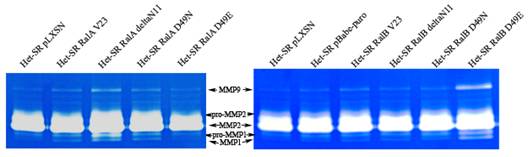
Рисунок 10. Желатиназная зимография кондиционированных сред, полученных от клеточных линий, экспрессирующих различные формы Ral
Влияние белков Ral на внутриклеточную передачу сигнала неоспоримо. Исследование влияния экспрессии различных форм белков Ral на активацию белковых каскадов в клетках Het-SR показало, что фосфорилирование по 183 тирозину киназы JNK1 (MAPK8) повышено в клеточных линиях с активными формами белков Ral (см. рисунок 11). Эффекторные мутанты RalA deltaN11 и RalA D49E не обладали способностью стимулировать активацию JNK1, в то время как все эффекторные мутанты RalB не отличались от исходной полностью активной формы по этому параметру. Таким образом, можно сделать вывод о том, что для подавления стимуляции JNK1 недостаточно блокирования взаимодействия RalB с одним из трех исследованных белков-партнеров, в то время как взаимодействие RalA с PLD и комплексом экзоциста являются необходимыми условиями для RalA-зависимой активации этой киназы. Нами также выявлено некоторое влияние белков Ral на активацию Stat3 и p38, но не ERK ½ (данные не приведены).

Рисунок 11. Вестерн-блот анализ уровня экспрессии P-JNK1
Кавеолин-1 (Cav-1) – основной структурный компонент кавеол, в которых концентрируются многие сигнальные молекулы. Cav-1 регулирует активность таких значимых в канцерогенезе белков, как Ha-Ras, Src, рецепторов факторов роста и многих других. Изменение экспрессии этого белка в опухолях и клеточных линиях интенсивно изучается, и есть данные, указывающие на то, что в подавляющем большинстве злокачественных новообразований снижение количества Cav-1 кореллирует с высокой степенью злокачественности опухолей. Однако, существует гипотеза, что экспрессия Cav-1 может носить цикличный характер и повышаться при метастазировании опухолей.
Cav-1 экспрессируется в исследованных линиях на очень высоком уровне. В клеточных линиях Het-SR, несущих эффекторные мутанты RalA deltaN11, RalB V23, RalB D49N и RalB D49E наблюдается слабовыраженное снижение экспрессии этого белка. Некоторое повышение его экспрессии отмечено в линии с активной формой RalA, а также в клетках с заблокированным взаимодействием RalA с комплексом экзоциста (см рисунок 12).

Рисунок 12. Вестерн-блот анализ экспрессии Кавеолина-1 (в качестве контроля нанесения использовались антитела к β-актину)
5. Анализ полученных данных
Очевидно, что усиление метастатического фенотипа в клетках может достигаться разными путями, так как способность формировать вторичные очаги роста – это комплексная характеристика. Опираясь на результаты нашего исследования (см. таблицу 1) можно сказать, что экспрессия активной формы RalB вносит больший вклад в усиление злокачественного фенотипа in vitro, чем ГТФ-связанная форма RalA. Более того, экспрессия RalB V23 приводит к усилению спонтанной метастатической активности клеток Het-SR, а также снижает минимальную прививочную дозу in vivo.
Одним из важных, с нашей точки зрения, наблюдений явилось значимое усиление СМА при экспрессии эффекторного мутанта RalA D49E, неспособного связываться с комплексом экзоциста. По характеристикам, ассоциированным с агрессивным фенотипом in vitro, эта клеточная линия оказалась в целом менее злокачественной, чем культура клеток с полностью активной формой RalA V23. Однако экспрессия формы RalA, неспособной связываться с Sec5, приводила к усилению инвазии через матригель и в ней повышалась экспрессия белка Cav-1.
C другой стороны, эффекторный мутант RalB deltaN11, неспособный взаимодействовать c PLD, по своему влиянию на клеточные характеристики in vitro отличался от полностью активной формы RalB лишь по некоторым характеристикам (снижал способность к неприкрепленному росту и миграции, не оказывал влияния на экспрессию Cav-1), но не обладал способностью усиливать СМА клеточной линии Het-SR.
Таблица 1. Сравнительный анализ изменения клеточных характеристик in vitro.
Het-SR RalA V23 | Het-SR RalA deltaN11 | Het-SR RalA D49N | Het-SR RalA D49E | Het-SR RalB V23 | Het-SR RalB deltaN11 | Het-SR RalB D49N | Het-SR RalB D49E | |
Метастазирование | (+) | (+) | (+) | (+++) | (+++) | (++) | (++) | |
Подвижность | (++) | (+) | (+) | |||||
Динамика пролиферации | (++) | (+) | (++) | (++) | (++) | (++) | ||
Формирование колоний в условиях разреженной популяции | (+) | (+) | (++) | (++) | (+) | (+) | (+) | |
Неприкрепленный рост | (-) | (-) | (-) | (-) | ||||
Инвазия in vitro | (+) | (++) | (++) | (+++) | (++) | (++) | ||
Подвижность по градиенту ростовых факторов | (+) | (+) | (+++) | (++) | (+) | (+) | ||
Секреция MMP | (+) | (+) | ||||||
Секреция uPA | (+) | (+) | ||||||
Фосфорилирование JNK1 | (+) | (+) | (+) | (+) | (+) | (+) | ||
Экспрессия CyclinD1 | (+) | (+) | (+) | (+) | ||||
Экспрессия CD24 | (+) | |||||||
Экспрессия Cav-1 | (+) | (-) | (++) | (-) | (-) | (-) | ||
Туморогенность | (-) | (+) | (+) | |||||
Размер опухолей | (+) | (+) | (+) | (-) | (-) | |||
(+) | (++) | (+++) | (-) | |||||
незначительное усиление по сравнению с контролем | усиление по сравнению с контролем | значительное усиление по сравнению с контролем | снижение по сравнению с контролем | признак не изменился по сравнению с контролем |
Таким образом, можно утверждать, что усиление спонтанной метастатической активности в данной модели - крайне сложный процесс, который не может быть предсказан с помощью одного или нескольких из использованных тестов. Однако участие белков Ral в этом процессе является неоспоримым. Более того, эти ГТФазы различаются по степени влияния на различные характеристики трансформированных клеток не только между собой, но и по набору эффекторов (PLD, RalBP1 и комплекс экзоциста), необходимых для осуществления этих функций.
Выводы
1. Получены линии, стабильно экспрессирующие различные формы белков RalA и RalB: активные ГТФ-связанные белки, а также эффекторные мутанты, неспособные взаимодействовать с PLD, RalBP1 или с комплексом экзоциста.
2. Активность белков Ral (в большей степени RalB) приводит к стимуляции спонтанной метастатической активности клеток Het-SR. Мутация, блокирующия взаимодействие с PLD, отменяет RalB-опосредованную стимуляцию метастазирования.
3. Белки Ral стимулируют пролиферацию, подвижность и рост в условиях разреженной популяции исследованных клеток.
4. Активная форма RalB в большей степени, чем RalA активирует инвазию in vitro, что коррелирует со стимуляцией миграции клеток по градиенту ростовых факторов. При этом для стимуляции инвазии как RalB, так и RalA необходимо взаимодействие с RalBP1.
5. Различные мутантые формы белков Ral влияют на активность белков, ремоделирующих ВКМ (MMP1 и MMP9). RalA, в отличие от RalB, усиливает активность протеазы uPA, что опосредовано взаимодейстивем с RalBP1 и комплексом экзоциста.
6. Активные формы RalA и RalB, а также их эффекторные мутанты по-разному модулируют внутриклеточную передачу сигнала и экспрессию белков, потенциально ассоциированных с метастазированием: фосфорилирование киназы JNK1, экспессию CyclinD1, Cav-1 и CD24.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
1. , , Активность протеиназ внеклеточного матрикса в экспериментальной модели опухолевой прогрессии.// Вестник РОНЦ. – 2008 – т.19., №4– с. 16-21.
2. , , Модуляторы активности фосфолипазы D белки RalA и Arf6: влияние на различные характеристики трансформированных клеток.// Цитология. – 2008 - т.50, №9 - с. 807.
3. Tchevkina E.M., Knizhnik A.V., Rybko V.A., Kainov Y.A., Komelkov A.V. Extracellular Proteases Activity In Tumor Progression Experimental Model.// Аnticancer Research. – 2, 5C - 3513.
4. Knizhnik A. V., Rybko V. A., Kainov Y. A., Komelkov A. V. and Tchevkina E. M. ARF6 and EСМ protease activity in hamster tumor progression experimental model.// Cancer Degradome Symposium, London, UK - 8-9 October 2008.
5. Зборовская И. Б., , Молекулярные маркеры метастазирования рака легкого в лаборатории и клинике.// «СПИД, рак и общественное здоровье»г. - Т.1
6. Knizhnik A., Rybko V., Aushev V., Kainov Y., Komelkov A., Tchevkina E. Arf6 participates in modulation of cell metastatic activity in vivo.// European Small GTPase meeting, 29-31 August, 2007, Umea, Sweden.
7. Rybko V., Ukraintsev K., Martinjuk A., Dyakova N., Komelkov A., Kovaleva O., Tchevkina E. RalA and RalB proteins contribute to metastasis stimulation through distinct pathways.// 2nd International Congress on Molecular Staging of Cancer, Mannheim/Heidelberg, - European Journal of Cancer Supplements.- 2006 - Volume 4, Issue
8. , , Экспрессия Кавеолина-1 в опухолях мягких тканей
|
