
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Вероятно, существует обратная зависимость скорости распластывания от активности миозина II. Во-первых, актин-миозиновое натяжение в пучках может сдерживать распластывание. Во-вторых, в присутствии Y27632 и блеббистатина может усиливаться полимеризация актиновой сети и протрузия края, например, за счет активации Rac.
2. Эффект ингибиторов сократимости на организацию и локомоторное поведение трансформированных фибробластов.
2.1. Изменения морфологиии фибробластов при RAS-трансформации.
При RAS-трансформации клетки становились хуже распластанными и образовывали хвосты и отростки в разных направлениях. Увеличивалось количество ровных вогнутых краев, уменьшались размеры ламелл. Другим характерным признаком RAS-трансформированных клеток было ослабление и утрата упорядоченной организации актин-миозиновых пучков. Фокальные контакты становились более мелкими. Трансформированные фибробласты имели более одного активного края, и участки псевдоподиальной активности были непостоянны. Так, в норме кратковременная активация Ras под воздействием внешнего стимула инициирует полимеризацию актина и формирование активного края и тем самым регулирует полярность клетки и направленность движения (Burridge and Wennerberg, 2004; Sasaki et al., 2004). Мы предполагаем, что при постоянной активации Ras в трансформированных клетках индукция полимеризации актина происходит спонтанно на разных участках края вне зависимости от каких-либо стимулов.
2.2. Морфология трансформированных клеток, обработанных ингибиторами сократимости.
При обработке Y27632 или блеббистатином трансформированные фибробласты образовывали длинные ветвящиеся отростки и тонкие хвосты, приобретая причудливую форму. Как и в случае действия ингибиторов на нетрансформированные клетки, актин-миозиновые пучки пропадали, в ламеллах выявлялась густая сеть микрофиламентов и интенсивное образование раффлов. Немногочисленные фокальные контакты исчезали, но оставались точечные фокальные комплексы.
Итак, при RAS-трансформации ослабевает система актин-миозиновых пучков и фокальных контактов. Действие ингибиторов сократимости на трансформированные клетки проявляется еще более полной утратой этих структур и образованием многочисленных отростков, имеющих псевдоподиальную активность.
2.3. Миграция трансформированных клеток, обработанных ингибиторами сократимости.
Трансформированные клетки мигрировали в рану более эффективно: их количество (N) и пройденные дистанции (L) всегда были больше, чем для контрольных нетрансформированных клеток. Было отмечено, что кривая миграции трансформированных фибробластов практически совпадает с кривой для нетрансформированных клеток, обработанных ингибиторами сократимости. Однако движение трансформированных клеток утрачивало направленный характер. Ненаправленный характер миграции в сочетании с высокой скоростью обусловлен, видимо, спонтанным и интенсивным образованием протрузий в трансформированных клетках.
При подавлении сократительной активности миозина II в трансформированных клетках дальность миграции также увеличивалась, но, как правило, дальше выходили лишь единичные клетки. Так, расстояние более 900 мкм преодолели менее 3% клеток. Общее же число мигрировавших трансформированных клеток в серии опытов не сильно различалось в присутствии и в отсутствии ингибиторов сократимости (табл. 4, рис. 4).
Табл. 4. Количество клеток и пройденные расстояния при миграции в рану трансформированных фибробластов, обработанных Y27632 и блеббистатином.
RAS | RAS+Y27632 | RAS+блебб. | |
N кл. | 436 | 372 | 441 |
100% | 85% | 101% | |
L, мкм | 900 | 1200 | 1300 |
100% | 133% | 144% |
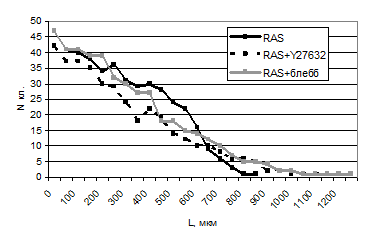 Рис. 4. Распределение по расстояниям при миграции в рану RAS-трансформированных клеток в присутствии блеббистатина и Y27632 (по результатам одного из 3 аналогичных экспериментов).
Рис. 4. Распределение по расстояниям при миграции в рану RAS-трансформированных клеток в присутствии блеббистатина и Y27632 (по результатам одного из 3 аналогичных экспериментов).
Мы увидели, что при RAS-трансформации подвижность фибробластов повышается, как и при подавлении сократимости в нетрансформированных клетках. Миграционная способность трансформированных клеток при воздействии ингибиторов миозина увеличивается в меньшей степени, нежели миграция нетрансформированных.
2.4. Эффект ингибиторов миозина II на динамику распластывания и поляризацию трансформированных фибробластов.
Распластывание трансформированных клеток происходило фрагментарно: псевдоподиальная активность не распределялась равномерно по всему краю, а сосредотачивалась на нескольких участках, где сразу начиналось образование отростков, таким образом, отсутствовала стадия радиального распластывания (рис. 3). На каждой стадии площадь клеток была примерно в два раза меньше, чем в контроле (табл. 3 и 5). Трансформированные клетки поляризовались раньше, и их степень поляризации была выше, чем у контрольных фибробластов, за счет повышения дисперсии, т. е. за счет образования отростков (табл. 3 и 5).
Под воздействием ингибиторов миозина скорость распластывания трансформированных клеток увеличивалась в меньшей степени, нежели в случае нетрансформированных, а степень поляризации (дисперсия) сильно возрастала за счет интенсивного образования хвостов и отростков (табл. 5).
площадь, кв. мкм | дисперсия | элонгация | степень поляризации | |
RAS 2ч | 844±45 | 0.140±0.023 | 0.499±0.057 | 0.639±0.067 |
4ч | 1159±65 | 0.606±0.056 | 0.943±0.087 | 1.548±0.116 |
24ч | 2283±146 | 0.808±0.081 | 1.306±0.114 | 2.115±0.114 |
RAS+Y27632 2ч | 1046±81 | 0.978±0.075 | 0.91±0.103 | 1.887±0.139 |
4ч | 1116±59 | 1.48±0.099 | 1.103±0.101 | 2.583±0.103 |
24ч | 2932±193 | 2.214±0.146 | 2.08±0.148 | 4.294±0.146 |
RAS+блебб. 2ч | 1216±53 | 1.043±0.078 | 0.86±0.103 | 1.903±0.107 |
4ч | 1794±80 | 0.96±0.092 | 1.154±0.142 | 2.114±0.185 |
24ч | 2793±146 | 2.344±0.155 | 1.921±0.145 | 4.265±0.144 |
Табл. 5. Данные морфометрического анализа контуров трансформированных клеток через 2, 4 и 24 ч распластывания при действии Y27632 и блеббистатина.
Итак, при сравнении фенотипов трансформированных клеток с нормальными клетками, обработанными ингибиторами миозина, мы выявили несколько общих черт: 1) ослабление актин-миозиновых структур и контактов; 2) повышение псевдоподиальной активности и усиление миграции; 3) увеличение степени поляризации. Отличия же состоят в том, что трансформированные клетки гораздо хуже распластаны и мигрируют менее направленно. Ухудшение распластывания в данном случае нельзя объяснить ослаблением актин-миозиновой системы, поскольку ингибирование миозина в нормальных клетках приводит к усилению распластывания. Мы полагаем, что при трансформации нарушения затрагивают систему адгезии (например, через активность белка Src), что и влечет «недоразвитие» системы стресс-фибрилл: мощные пучки не могут развиться на слабых контактах. Во-первых, это объясняет тот факт, что эффект ингибиторов миозина становится менее выражен. Если ослабление пучков при трансформации вызвано ухудшением адгезии, а уровень активации миозина сохраняется, то ингибитор будет одинаково снижать ее в нормальных и трансформированных клетках. А так как пучки трансформированных клеток были слабее, то и видимый эффект ингибитора будет казаться меньше. Во-вторых, ослабленная система контактов не может поддерживать распластанное состояние, поскольку натяжение пучков ведет к легкому отрыву и подтягиванию клеточных краев. Поэтому площадь трансформированных клеток снижается в два раза.
3. Ультраструктура цитоскелета клеток при разной степени подавления миозиновой сократимости.
3.1. Ультраструктура нетрансформированных фибробластов.
На электронно-микроскопическом уровне, также как и на световом, выявлялись мощные актиновые пучки, закрепленные на крупных фокальных контактах. Клетки формировали плоские ламеллиподии, образованные густой актиновой сетью. Для определения локализации миозина мы использовали антитела, конъюгированные с коллоидным золотом. Миозин II был колокализован с актиновыми пучками. В контрольных клетках филаменты миозина располагались упорядоченно и формировали регулярные структуры в виде цепочек и «стеков» (рис. 5).
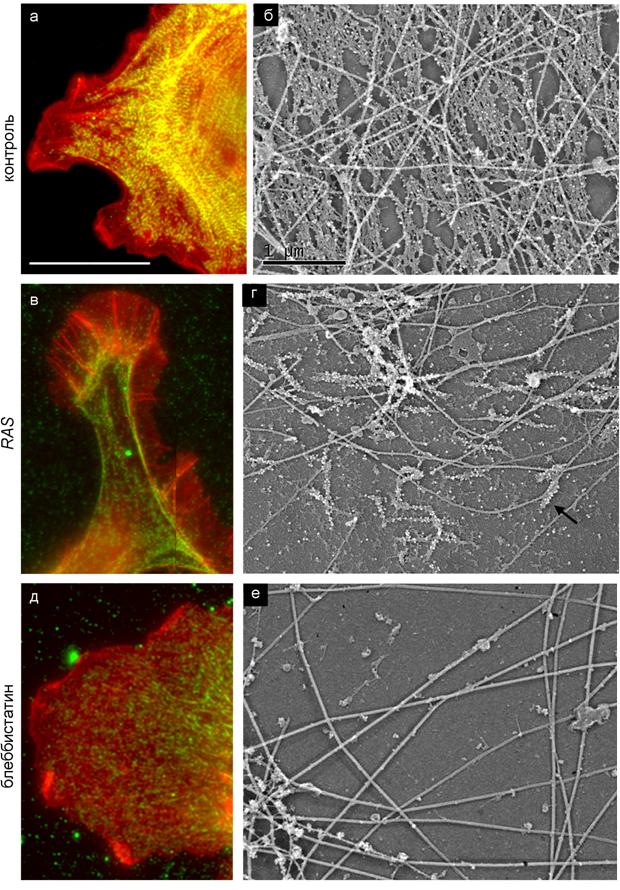 Рис. 5. Организация миозина в нормальных, трансформированных и обработанных блеббистатином клетках. (а, в, д) Флуоресцентная окраска на актин (красный) и миозин II (зеленый) Масштаб 20 мкм. (б, г, е) Платиновые реплики цитоскелета (актин удален с помощью гельзолина). Стрелкой указан миозиновый филамент.
Рис. 5. Организация миозина в нормальных, трансформированных и обработанных блеббистатином клетках. (а, в, д) Флуоресцентная окраска на актин (красный) и миозин II (зеленый) Масштаб 20 мкм. (б, г, е) Платиновые реплики цитоскелета (актин удален с помощью гельзолина). Стрелкой указан миозиновый филамент.
3.2. Ультраструктура клеток, обработанных блеббистатином.
Клетки, обработанные 45-75 мкМ блеббистатина, утрачивали мощные стресс-фибриллы, связанные со зрелыми фокальными контактами, что приводило к общему расслаблению клетки и неспособности к ретракции. Такие клетки хорошо прикрепляются к субстрату, образуют ламеллиподии и имеют высокую подвижность. На электронно-микроскопическом уровне мы обнаруживали тонкие актин-миозиновые пучки в центральной части ламеллы и равномерную сеть актина лишь непосредственно позади ламеллиподий. Мы решили посмотреть, как будет меняться структура ламеллы при более глубокой степени ингибирования миозина, и использовали также блеббистатин в концентрации 100 мкМ. Клетки, обработанные 100 мкМ блеббистатина, становились поджатыми, ламеллиподии пропадали, а фокальные комплексы выявлялись редко. Вместо ламеллиподий мы наблюдали образование филоподий и раффлов. Мы увидели, что большая часть миозина переходит в несвязанное с актином состояние и резко снижается количество миозиновых филаментов (рис. 5). К настоящему моменту не была показана зависимость сборки биполярных филаментов от АТФ-азной активности миозина II.
Мы заключили, что протрузия (образование ламеллиподий) и прикрепление края (образование фокальных комплексов) взаимозависимы, а для эффективного прикрепления необходима минимальная активность миозина II.
По традиционным представлениям, миозин-зависимое сокращение обеспечивает построение пучков и созревание контактов, но не протрузию ведущего края. Полимеризация актина в ламеллиподии и появление фокальных комплексов считается независимым от натяжения. Вероятно, инициация сборки фокальных комплексов действительно происходит без участия миозина, но они быстро подвергаются разборке, если к ним не приложено механическое натяжение. По нашим наблюдениям, ламеллиподии успешно формируются, только если они могут отталкиваться от сайтов адгезии, таким образом, они косвенно зависят от контрактильности миозина.
3.3. Ультраструктура RAS-трансформированных клеток.
Мы показали, что в трансформированных фибробластах также как и в контрольных клетках миозин организован в биполярные филаменты. Однако они не имеют единой ориентации и гораздо реже формируют регулярные структуры (рис. 5). При обработке трансформированных клеток блеббистатином миозиновые филаменты практически исчезают.
4. Восстановление актин-миозиновых структур при отмывке ингибиторов миозина II.
Изменения структуры цитоскелета под действием Y27632 и блеббистатина были полностью обратимы. Мы наблюдали очень быстрое (в течение минут) восстановление актин-миозиновых структур при отмывке от 100 мкМ блеббистатина. Мы показали, что первым видимым изменением является образование и прикрепление ламеллиподий (мощные ламеллиподии и фокальные комплексы в их основании развиваются в течение 1 мин отмывки). Последовательно повышается фракция связанного с цитоскелетом миозина и степень агрегации его в филаменты, в течение 5-15 мин развиваются стресс-фибриллы и созревают фокальные контакты.
5. Эффект ингибиторов полимеризации актина на локомоторное поведение нетрансформированных и трансформированных клеток.
5.1. Морфология клеток, обработанных ингибиторами полимеризации актина.
Мы подобрали низкие концентрации агентов, которые не ингибировали полимеризацию актина полностью, а лишь снижали ее интенсивность. При действии 0.5 мкМ латрункулина А, нетрансформированные клетки напоминали по форме контрольные, однако они становились более поджатыми. Клетки формировали мощные актин-миозиновые пучки, но ламеллиподии исчезали (рис. 3). Сохранялись только зрелые фокальные контакты на концах стресс-фибрилл, а новых фокальных комплексов не образовывалось. Действие цитохалазина D проявлялось сходным образом. Действие ингибиторов полимеризации актина на трансформированные фибробласты проявлялось абсолютно иначе: клетки формировали аномальные ламеллиподии и мелкие контакты при сильной дезорганизации остального актинового цитоскелета (рис. 3). Видеонаблюдение показало, что под действием латрункулина такие клетки сохраняли псевдоподиальную активность.
Мы показали, что низкие концентрации ингибиторов полимеризации актина подавляли в первую очередь образование ламеллиподий, но не пучков в нормальных клетках, тогда как в трансформированных клетках практически исчезали пучки и сохранялись ламеллиподии.
5.2. Миграция клеток, обработанных ингибиторами полимеризации актина.
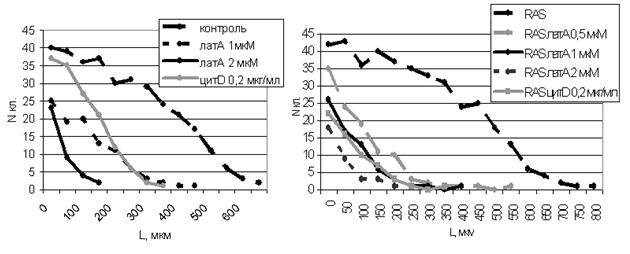
Воздействие низких доз латрункулина А и цитохалазина D приводило к значительному подавлению миграционной способности как нормальных так и трансформированных клеток (табл. 6 и 7, рис. 6).
Табл. 6. Количество клеток и пройденные расстояния при миграции в рану контрольных клеток и обработанных латрункулином А и цитохалазином D.
контр. | лат. A 1мкМ | лат. A 2мкМ | цит. D | |
N кл. | 326 | 101 | 38 | 141 |
100% | 31% | 12% | 43% | |
L, мкм | 700 | 500 | 200 | 400 |
100% | 71% | 29% | 57% |
Табл. 7. Количество клеток и пройденные расстояния при миграции в рану трансформированных фибробластов, обработанных латрункулином А и цитохалазином D.
RAS | RAS+лат. A 0.5 мкМ | RAS+лат. A 1 мкМ | RAS+лат. A 2 мкМ | RAS+цит. D | |
N кл. | 391 | 108 | 68 | 35 | 60 |
100% | 28% | 17% | 9% | 15% | |
L, мкм | 850 | 600 | 450 | 300 | 400 |
100% | 71% | 53% | 35% | 47% |

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
