
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ШУТОВА Мария Сергеевна
РОЛЬ АКТИН-МИОЗИНОВЫХ СТРУКТУР В МЕХАНИЗМАХ ПОДВИЖНОСТИ НОРМАЛЬНЫХ И ТРАНСФОРМИРОВАННЫХ ФИБРОБЛАСТОВ
Специальность 14.01.12 – Онкология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Москва 2010
Работа выполнена в лаборатории механизмов канцерогенеза НИИ канцерогенеза Учреждения Российской Академии Медицинских Наук Российского онкологического научного центра им. РАМН.
Научный руководитель: доктор медицинских наук
член-корр. РАН, профессор
ВАСИЛЬЕВ Юрий Маркович
Официальные оппоненты: доктор медицинских наук, профессор
СТАВРОВСКАЯ Алла Александровна
доктор биологических наук, профессор
СМИРНОВА Елена Александровна
Ведущая организация: Институт цитологии
Российской Академии Наук
Защита диссертации состоится « » 2010 г. в часов на заседании диссертационного совета (Д.001.017.01) Учреждения Российской Академии Медицинских Наук Российского онкологического научного центра имени РАМН.
С диссертацией можно ознакомиться в библиотеке Учреждения Российской Академии Медицинских Наук Российского онкологического научного центра имени РАМН.
Автореферат разослан «___»_____________ 2010 года.
Ученый секретарь
диссертационного совета, доктор медицинских наук, профессор
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы
При опухолевой прогрессии нарушаются не только механизмы нормальной пролиферации клеток, но и их локомоторная активность, что приводит к таким проявлениям «асоциального» поведения опухолевых клеток, как инвазивный рост и метастазирование.
На клеточном уровне основой возникновения этих свойств являются генетические изменения, приводящие к нарушению регуляции адгезии и подвижности клеток (Yamazaki et al., 2005). Установлено, что в этих процессах определяющую роль играют перестройки цитоскелетных структур. Трансформированные клетки зачастую отличаются от нормальных по своей морфологии и организации цитоскелета (Bershadsky and Vasiliev, 1988). Реорганизации цитоскелета, особенно изменения клеточной сократимости, регулируемой актин-миозиновым комплексом, имеют центральное значение для развития фенотипа морфологически трансформированных клеток с инвазивным поведением. Редукция стресс-фибрилл, характерная для многих типов трансформированных клеток, сопряжена с нарушением созревания контактных структур (Ровенский и Васильев, 2004) и часто коррелирует с повышением локомоторной активности и/или метастатического потенциала опухолевых клеток (Pokorna et al., 1994; Sahai and Marshall, 2003).
Таким образом, изучение механизмов нормальной подвижности клетки и нарушений этих механизмов является одной из важнейших задач современной клеточной биологии.
Согласно современным данным, главную роль в определении локомоторного фенотипа играет динамическое взаимодействие двух основных систем цитоскелета – микротрубочек (МТ) и актин-миозиновой системы, создающей внутри клетки и в окружающем ее матриксе механическое натяжение, регулируемое молекулярной системой малых ГТФ-аз семейства Rho. Предыдущие работы нашей лаборатории дают основание предположить, что одним из ключевых событий в приобретении способности к инвазии является Rho-зависимое снижение актин-миозиновой контрактильности при сохранении структуры и активности системы МТ. Заслуживает внимания дальнейшее исследование механизмов Rhо-зависимой регуляции реорганизаций цитоскелета, а также формы, адгезии и движения в одном из основных морфологических типов клеток – в фибробластах.
Цель и задачи исследования
Целью работы является изучение роли актин-миозиновой системы в определении морфологии и подвижности фибробластов в норме и при RAS-трансформации. Мы использовали клетки линии REF52tetRas с тетрациклин-регулируемой экспрессией онкогена RAS, в которых отмывка тетрациклина приводит к синтезу конститутивно активного белка и к морфологической трансформации клеток (Kopnin et al., 2003; Alexandrova et al., 2006).
Задачи исследования:
1. Охарактеризовать изменения морфологии, подвижности и направленности движения нетрансформированных фибробластов при подавлении актин-миозиновой сократимости (с помощью ингибиторов Y27632 и блеббистатина) по следующим пунктам:
- исследование строения цитоскелетных структур и фокальных контактов иммунофлуоресцентными методами;
- видеонаблюдение с целью изучения поведения индивидуальных клеток;
- оценка скорости и степени распластывания клеток с помощью компьютерного морфометрического анализа;
- оценка эффективности направленной миграции клеток в экспериментальную рану;
- ультраструктурное исследование цитоскелета методом платиновых реплик;
- исследование динамики восстановления актин-миозиновых структур после отмывки ингибиторов сократимости.
2. Охарактеризовать изменения в морфологии и подвижности фибробластов при RAS-трансформации.
3. Сравнить изменения в морфологии и локомоторном поведении трансформированных и нетрансформированных клеток в присутствии агентов, изменяющих организацию актин-миозинового цитоскелета (Y27632, блеббистатин, латрункулин А, цитохалазин D).
Научная новизна и практическая значимость работы
Необходимость дальнейших исследований молекулярных и клеточных механизмов морфологической трансформации и инвазивных миграций опухолевых клеток не подлежит сомнению. Взаимодействие различных компонентов цитоскелета и регуляторные пути при развитии инвазивного фенотипа еще не достаточно изучены.
В настоящей работе впервые проведен детальный анализ динамики актин-миозинового цитоскелета и локомоторного поведения трансформированных фибробластов в сравнении с нетрансформированными, а также проанализирована роль двух основных процессов – полимеризации актина и актин-миозинового сокращения – в клеточной подвижности.
В последние годы получены новые соединения, избирательно меняющие активность белков, регулирующих динамику цитоскелета, это Y27632, ингибитор активирующей миозин Rho-киназы (Ishizaki et al., 2000), и блеббистатин, ингибитор немышечного миозина II (Straight et al., 2003). В нашей работе впервые проведено сравнение локомоторных реакций нормальных и трансформированных фибробластов на эти агенты, а также на ингибиторы полимериации актина (латрункулин А и цитохалазин D).
Мы показали, что трансформированные клетки имеют сходство с клетками, обработанными ингибиторами сократимости: это усиление локомоторной активности на фоне ослабления/разрушения системы актин-миозиновых пучков. Однако, в отличие от трансформированных клеток, нормальные фибробласты сохраняют способность к направленному движению и под действием ингибиторов сократимости, которое поддерживается системой МТ.
Показано, что трансформированные клетки иначе реагируют на низкие концентрации ингибиторов полимеризации актина, а именно утрачивают актин-миозиновые пучки, но не псевдоподиальную активность. Напротив, нормальные клетки утрачивают активность края, но сохраняют пучки.
Впервые показано, что минимальная активность немышечного миозина II играет ключевую роль в первичном прикреплении и эффективной протрузии клеточного края. Также показано, что организация миозина в «рабочие единицы» – биполярные филаменты – зависит от степени и характера развития стресс-фибрилл в клетке.
Таким образом, ослабление актин-миозиновой системы в значительной мере обуславливает усиление подвижности RAS-трансформированных клеток, хотя, вероятно, ненаправленность движения обусловлена активацией других сигнальных путей, ведущих от онкобелка Ras к цитоскелету.
Изучение механизмов регуляции перестроек цитоскелета и подвижности клеток, возможно, в дальнейшем откроет подходы к направленному исправлению молекулярных и клеточных нарушений, приводящих к инвазии, например, через нормализацию функций регуляторных белков, в частности, Rho-системы.
Апробация работы
Диссертация апробирована на совместной научной конференции лабораторий механизмов канцерогенеза, механизмов прогрессии эпителиальных опухолей, иммунохимии, генетики опухолевых клеток НИИ канцерогенеза РОНЦ им. РАМН и отдела математических методов в биологии НИИ физико-химической биологии им. МГУ 30 сентября 2009 г.
Материалы диссертационной работы были представлены на XII и XIII международных конференциях студентов, аспирантов и молодых ученых «Ломоносов» (Москва, 2005 и 2006); на конференции «Молекулярные механизмы процессов онтогенеза: эмбриогенез, геномы, эволюция» (Москва, 2006); на конференции «Биология клетки в культуре» (Санкт-Петербург, 2006); на ХII Российском онкологическом конгрессе (Москва, 2008); а также на международных конференциях The American Society for Cell Biology 48th Annual Meeting (San Francisco, 2008) и The American Society for Cell Biology 49th Annual Meeting (San Diego, 2009).
Публикации
По материалам диссертации опубликовано 10 работ.
Структура и объем работы
Диссертационная работа изложена на 144 страницах, содержит 64 рисунка и 9 таблиц и состоит из следующих разделов: Списка сокращений, Введения, Обзора литературы, Материалов и Методов, Результатов, Обсуждения, Выводов и Списка литературы, включающего 163 цитируемых источника.
МАТЕРИАЛЫ И МЕТОДЫ
Клеточная культура. В работе использовали клетки линии REF52tetRas, любезно предоставленные проф. (Agapova et al., 1999; Kopnin et al., 2003). Клетки были получены из исходной линии REF52 (иммортализованные крысиные эмбриональные фибробласты) путем трансфекции плазмидой с геном конститутивно активного белка N-Ras/asp12 под тетрациклин-зависимым супрессором. Таким образом, мы имели возможность регулировать экспрессию онкогена N-RAS и тем самым вызывать трансформацию клеток путем добавления/отмывки тетрациклина из среды. Клетки культивировали в среде DMEM (Sigma) с добавлением 10% эмбриональной сыворотки (PAA Laboratories GmbH) и антибиотиков (пенициллин, стрептомицин, 100 ед./мл) при 37оС и 5% СО2. Клетки вели в среде с добавлением тетрациклина (2 мкг/мл) каждые три дня. Для трансформации клеток тетрациклин отмывали и культивировали клетки в среде без тетрациклина в течение 5 дней, после чего использовали в экспериментах.
Ингибиторы. В экспериментах использовали следующие ингибиторы: Y27632 – ингибитор Rho-киназы (45 мкМ) (Calbiochem); блеббистатин – ингибитор АТФ-азной активности немышечного миозина II (45, 75 и 100 мкМ) (Toronto Research Chemicals Inc.); ингибиторы полимеризации актина – латрункулин А (0.5, 1, 2 мкМ) (Calbiochem) и цитохалазин D (0.2 мкг/мл) (Sigma); колцемид – агент, деполимеризующий микротрубочки (0.5 мкг/мл) (Sigma).
Иммунофлуоресцентное окрашивание и микроскопия. Для иммунофлуоресцентного окрашивания цитоскелетных структур применяли фиксацию 3.7% параформальдегидом на PBS, либо холодным метанолом (для окрашивания МТ). Для одновременного окрашивания МТ и микрофиламентов, клетки фиксировали 2.5% глютаральдегидом. Для окрашивания F-актина применяли флуоресцентные красители alexa 488 и 594 phalloidin (Molecular Probes). В исследовании были использованы следующие первые антитела: pAb rabbit-anti-Myosin (Non-muscle) IgG (Biomedical Technologies Inc.), pAb rabbit anti-p34-Arc/ARPC2 (Upstate, Millipore), mouse monoclonal anti-vinculin (Сlone hVIN-1, Sigma), anti-α-tubulin mouse monoclonal IgG1 (Clone DM1-A, Sigma), anti-acetylated tubulin mouse monoclonal IgG2B (Clone 6-11-B-1, Sigma), pAb rabbit anti-Phospho-Myosin Light Chain 2 (Thr18/Ser19) (Cell Signaling Technology). В качестве II антител использовали: TRITC goat anti-mouse (Sigma), TRITC goat anti-rabbit (Molecular Probes), alexa 488 goat anti-mouse (Molecular Probes), alexa 488 goat anti-rabbit (Molecular Probes), FITC goat anti-mouse IgG1 (Southern Biotech), TRITC goat anti-mouse IgG2B (Southern Biotech).
Препараты исследовали с помощью флуоресцентного микроскопа Axioplan (Zeiss) (20×, 40×, 100×), камеры Olympus DP70 c программным обеспечением DP Controller. Мы использовали также флуоресцентный микроскоп Eclipse Ti (Nikon) (100×) c камерой Hamamatsu ORCA ER (Hamamatsu Photonics) и программным обеспечением NIS-Elements AR.
Оценка скорости распластывания и морфометрический анализ. Для характеристики распластывания клетки (50 тыс. кл./мл) высевали в 35 мм-чашки Петри на покровные стекла и фиксировали через 2, 4 и 24 ч, затем окрашивали фаллоидином и фотографировали (40×). Полученные изображения одиночных клеток (30 клеток на точку) использовали для морфометрического анализа. Контуры клеток в цифровом виде были проанализированы с помощью программы TRACER V1.0 (Dunn and Brown, 1986). Скорость распластывания оценивали путем сравнения площади и степени поляризации клеток на разных сроках фиксации. Степень поляризации определяли как сумму двух параметров – дисперсии и элонгации клеточного контура (Brown et al., 1989).
Анализ миграции в рану и направленности движения. Миграционную способность оценивали по стандартной методике – анализу миграции клеток в экспериментальную рану (Valster et al., 2005). Клетки высаживали на размеченные сеткой покровные стекла 18×18 мм (Photoetched Coverslips, Bellco Biotechnology, Inc.), доращивали до монослоя и удаляли часть монослоя с помощью лезвия бритвы. Фотографировали 5 полей зрения (участков раны) на точку с использованием инвертированного микроскопа Axiovert 200 (Zeiss) (10×) и камеры AxioCam MRc (Zeiss) с программным обеспечением Axiovision 4.3. Далее клетки инкубировали в присутствии/отсутствии ингибиторов в течение суток. Через 24 ч после нанесения раны клетки фиксировали и окрашивали DAPI (Sigma). После чего по разметке на стеклах были найдены сфотографированные ранее поля зрения и вновь сфотографированы в режиме фазы и флуоресценции.
Количественную оценку миграции в рану осуществляли путем подсчета клеток, вышедших в рану на определенное расстояние за сутки на участке шириной 1000 мкм. Полученные кадры накладывали друг на друга в программе Adobe Photoshop CS2, что позволяло производить точный подсчет клеток, вышедших за изначальную границу раны. Пространство раны, начиная от бывшей границы, разбивали на полоски по 50 мкм и подсчитывали количество клеток в каждом. На основании полученных данных строили графики распределения клеток по расстояниям. Количество мигрировавших опытных клеток и максимальное расстояние выражали в процентах по отношению к контролю.
Для оценки направленности движения клеток фазовые изображения получали с интервалом в 1 ч в течение 4 ч через сутки после нанесения раны (30 клеток на точку). Позицию клетки определяли по положению ядра. Направленность миграции оценивали как отношение D/T (расстояние между начальной и конечной точками пути клетки, поделенное на общую длину пути). D, T и скорость миграции клеток были измерены с помощью программы ImageJ.
DIC-видеомикроскопия. Для видеонаблюдения клетки высаживали на покровные стекла площадью 18×18 мм. За 24 ч до начала видеосъемки наносили широкую рану в монослое. Ингибиторы добавляли перед видеосъемкой. Для видеонаблюдения использовали клетки, обработанные Y27632, но не блеббистатином, поскольку блеббистатин инактивируется под воздействием света (Kolega, 2004). DIC-видеомикроскопию проводили с использованием микроскопа Leitz Aristoplan (40×) в условиях постоянной температуры. Видеозапись проводили с интервалом 30 сек между кадрами в течение 2-4 ч с помощью цифровой камеры Hamamatsu (C8484-05G, Hamamatsu Photonics) с программным обеспечением Wasabi (Hamamatsu Photonics Deutschland GmbH).
Платиновые реплики и электронная микроскопия. Коррелятивная электронная микроскопия платиновых реплик цитоскелета позволяла одновременно рассмотреть взаимную пространственную организацию актиновых, миозиновых и контактных структур в одной клетке при большом разрешении электронного микроскопа. Метод получения платиновых реплик с цитоскелетных препаратов включает в себя следующие этапы обработки культивируемых клеток: экстракция, фиксация, высушивание методом перехода критической точки, напыление, отмывка реплик (подробно см. Svitkina and Borisy, 1998; Svitkina, 2007). Для получения флуоресцентных изображений клетки окрашивали после экстракции и фотографировали с помощью инвертированного микроскопа Eclipse TE2000 (Nikon) (100×), камеры Cascade 512B (Roper Scientific, Trenton, NJ) с программным обеспечением MetaMorph (Molecular Devices, Sunnyvale).
Для визуализации миозина на электронно-микроскопическом уровне клетки окрашивали вторыми антителами, конъюгированными с коллоидным золотом: 18 nm Colloidal gold donkey anti-rabbit (Immunoresearch Laboratories Inc.), 1:5 в буфере А с 1% BSA. Поскольку сеть актиновых филаментов в клетке перекрывает другие цитоскелетные структуры при исследовании реплик, для подробного рассмотрения миозиновых структур готовили препараты, где актин был удален с помощью разрезающего белка гельзолина (0.4 мкг/мл в MES-KOH pH 6.3).
Платиновые реплики цитоскелета исследовали с помощью трансмиссионного электронного микроскопа JEOL 1200EX и CCD камеры ORIUS 835.10W (Gatan) с программным обеспечением Gatan Digital Micrograph. Изображения представляли в инвертированном виде.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1. Эффект ингибиторов сократимости на организацию и локомоторное поведение нетрансформированных фибробластов.
1.1. Контрольные нетрансформированные клетки.
Клетки линии REF52tetRas, культивируемые в присутствии тетрациклина, морфологически не отличались от исходной линии крысиных эмбриональных фибробластов REF52. Они были хорошо распластаны и поляризованы, имели мощные стресс-фибриллы и крупные фокальные контакты. Миозин II имел четкую колокализацию с актиновыми пучками и давал так называемую поперечную исчерченность. Контрольные клетки были способны к направленной миграции в экспериментальную рану. В процессе движения они формировали широкую ламеллу на переднем крае и 1-2 коротких хвоста, которые периодически подтягивались.
1.2. Локомоторный фенотип клеток с подавленной сократимостью.
Для подавления клеточной сократимости мы использовали: Y27632 – ингибитор активирующей миозин Rho-киназы и блеббистатин – ингибитор АТФ-азной активности миозина II. Было показано, что 45 мкМ Y27632 и 45 мкМ блеббистатина вызывали сходные изменения. При этой концентрации ингибиторов в клетках пропадали мощные актин-миозиновые пучки и зрелые фокальные контакты. Вдоль клеточного края сохранялись ламеллиподии и точечные фокальные комплексы. Миозин II окрашивался диффузно, поперечная исчерченность пропадала. При проведении экстракции клеток перед фиксацией интенсивность окраски миозина существенно снижалась, т. е. большая часть миозина была не связана с актином и вымывалась.
Движение клеток, обработанных Y27632 (блеббистатин не использовали для видеонаблюдения), сопровождалось значительными изменениями клеточной формы. Наиболее значимым изменением было образование и удлинение несократимых хвостов на заднем конце клетки. Ламеллы расширялись, боковые стабильные края пропадали, и на их местах формировались новые протрузии. Боковые ламеллы периодически коллапсировали и превращались в хвосты. Но псевдоподиальная активность никогда не возникала в уже сформированных хвостах. Всегда существовала одна ведущая ламелла, за которой направленно двигалось тело клетки.
В клетках, обработанных Y27632 или блеббистатином, большая часть МТ была направлена к активному краю, а остальные формировали в хвостах плотные тяжи, обогащенные ацетилированным тубулином, что свидетельствовало о повышенной стабильности МТ.
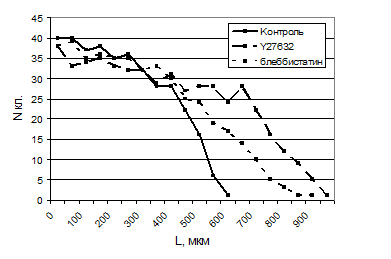 Эффективность миграции обработанных клеток значительно увеличивалась по сравнению с контролем: за 24 ч количество мигрировавших клеток возрастало приблизительно в полтора раза, и максимальная дальность миграции была также почти в полтора раза выше (табл. 1, рис. 1).
Эффективность миграции обработанных клеток значительно увеличивалась по сравнению с контролем: за 24 ч количество мигрировавших клеток возрастало приблизительно в полтора раза, и максимальная дальность миграции была также почти в полтора раза выше (табл. 1, рис. 1).
Рис. 1. Распределение клеток по расстояниям при миграции в рану в присутствии ингибиторов миозина II. Диаграмма иллюстрирует результаты одного из трех аналогичных опытов.
Табл. 1. Результаты анализа миграции контрольных и обработанных ингибиторами клеток. N кл. – общее число клеток, вышедших за границу раны за 24 ч; L – максимальное расстояние, пройденное клетками за 24 ч. Результаты для клеток с ингибиторами представлены как процент от контроля ± SEM.
контр. | Y27632 | блебб. | колц. | Y27632+колц | бл.+колц. | |
N кл., % | 100 | 150.6±12.6 | 146.9±26.6 | 18.8±9.8 | 17.0±8.0 | 16.6±4.8 |
L, % | 100 | 160.6±14.4 | 170.7±16.8 | 36.5±7.9 | 45.7±9.9 | 40.1±4.3 |
Наши данные показывают, что, вопреки распространенному мнению, миозин-зависимая ретракция не является необходимой для направленного клеточного движения: полярность клетки и способность к направленной миграции сохраняется в присутствии ингибиторов миозина II. С другой стороны, высокая степень развития стресс-фибрилл, видимо, не способствует высокой скорости перемещения: очень много актина занято в построении пучков и затруднена разборка старых фокальных контактов.
1.3. Способность клеток с подавленной сократимостью к поляризации.
Способность к поляризации мы оценивали по двум характеристикам – это способность клеток к направленному движению и способность к развитию поляризованного фенотипа в процессе распластывания.
Направленность миграции. В результате ингибирования миозина направленность движения не претерпевала изменений, а скорость значительно повышалась по сравнению с контролем (p<0.001 для Y27632 и блеббистатина согласно тесту Стьюдента) (табл. 2).
Табл. 2. Направленность и скорость движения нетрансформированных клеток при миграции в экспериментальную рану в течение 4 ч.
контроль | Y27632 | блеббистатин | |
Скорость, мкм/ч | 19.2±0.7 | 40±1.7 | 26.7±1.0 |
D/T | 0.90±0.02 | 0.95±0.01 | 0.94±0.01 |
Развитие поляризации de novo при распластывании. В процессе распластывания как контрольные, так и обработанные ингибиторами миозина клетки приобретали сначала дисковидную форму, а затем начинали поляризоваться. Поляризация контрольных клеток сопровождалась ориентацией стресс-фибрилл в едином направлении и последующим вытягиванием клетки. При ингибировании миозина первыми признаками поляризации клеток было с перераспределение МТ и псевдоподиальной активности. Через 4 ч в большинстве клеток был виден активный ведущий край, обогащенный микрофиламентами, и стабильный задний край, где утрачивалась псевдоподиальная активность и инициировались хвосты. К переднему активному краю клетки подходило множество МТ, тогда как сзади их было значительно меньше.
1.4. Потеря способности к поляризации и направленному движению при деполимеризации МТ колцемидом.
Один из возможных механизмов участия системы МТ в организации направленного движения подразумевает взаимодействие МТ с актин-миозиновыми пучками и фокальными контактами (Small and Kaverina, 2003). Мы показали, что при ингибировании миозина и утрате мощных пучков и контактов нетрансформированные клетки сохраняют способность к поляризации и направленному движению. Клетки формировали хвосты, содержащие тяжи стабильных МТ, и эти хвосты всегда находились на заднем конце мигрирующей клетки, а начало развития поляризации при распластывании совпадало с перераспределением МТ. Мы решили проверить, действительно ли система МТ регулирует направленное движение клетки в отсутствие стресс-фибрилл.
Разборка МТ колцемидом приводила к потере поляризации контрольных клеток. Мигрирующие в рану клетки утрачивали направление движения и останавливались в течение 10-20 мин после добавления колцемида, несмотря на высокую псевдоподиальную активность. Соответственно, миграция в рану была сильно подавлена (табл. 1, рис. 2).
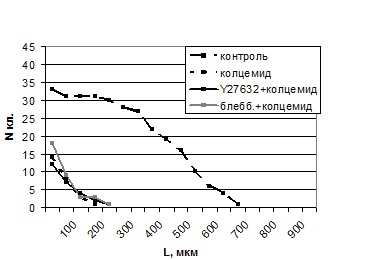 При добавлении колцемида к клеткам, предобработанным Y27632 или блеббистатином, большая часть МТ исчезала, но стабильные МТ часто сохранялись в хвостах. Однако клетки также утрачивали поляризацию и способность к направленному движению (табл. 1, рис. 2). Таким образом, ингибиторы сократимости никак не увеличивали скорость миграции в присутствии колцемида.
При добавлении колцемида к клеткам, предобработанным Y27632 или блеббистатином, большая часть МТ исчезала, но стабильные МТ часто сохранялись в хвостах. Однако клетки также утрачивали поляризацию и способность к направленному движению (табл. 1, рис. 2). Таким образом, ингибиторы сократимости никак не увеличивали скорость миграции в присутствии колцемида.
Рис. 2. Колцемид ингибирует миграцию в рану как необработанных, так и обработанных Y27632 или блеббистатином клеток. Диаграмма иллюстрирует один репрезентативный эксперимент.
Исходя из этих данных, мы предположили, что направленность движения клеток с подавленной активностью миозина определяется упорядоченным пространственным распределением МТ в интегрированной системе, т. е. относительным положением тяжа МТ в хвосте и МТ в ведущей ламелле. Возможно, собранные в пучок стабильные МТ хвоста не способствуют полимеризации актина и протрузии края, тогда как полимеризация поддерживается и усиливается МТ в ламелле.
1.5. Клетки с пониженной сократимостью распластываются быстрее.
Одним из проявлений клеточной подвижности является способность клетки к распластыванию. Мы исследовали динамику распластывания фибробластов через 2, 4 и 24 ч после посадки. Мы показали, что фибробласты с подавленной сократимостью распластывались интенсивнее, т. е. быстрее достигали конечной площади, чем контрольные клетки, и имели более высокую степень поляризации за счет образования длинных хвостов, тогда как конечная площадь обработанных клеток практически не отличалась от контроля (табл. 3, рис. 3).
Табл. 3. Данные морфометрического анализа контуров клеток через 2, 4 и 24 ч распластывания в контроле и при обработке Y27632 и блеббистатином.
площадь, кв. мкм | дисперсия | элонгация | степень поляризации | |
контроль 2ч | 1551±82 | 0.035±0.005 | 0.256±0.03 | 0.291±0.033 |
4ч | 2541±92 | 0.048±0.007 | 0.521±0.064 | 0.569±0.067 |
24ч | 4665±209 | 0.177±0.022 | 1.178±0.05 | 1.355±0.053 |
Y27632 2ч | 2860±94 | 0.013±0.001 | 0.211±0.023 | 0.224±0.024 |
4ч | 3213±105 | 0.07±0.011 | 0.387±0.052 | 0.457±0.059 |
24ч | 4689±247 | 1.361±0.146 | 1.445±0.131 | 2.805±0.212 |
блебб. 2ч | 2180±96 | 0.045±0.006 | 0.43±0.053 | 0.476±0.056 |
4ч | 2849±147 | 0.098±0.011 | 0.456±0.058 | 0.553±0.061 |
24ч | 5566±322 | 1.774±0.131 | 1.574±0.119 | 3.348±0.199 |
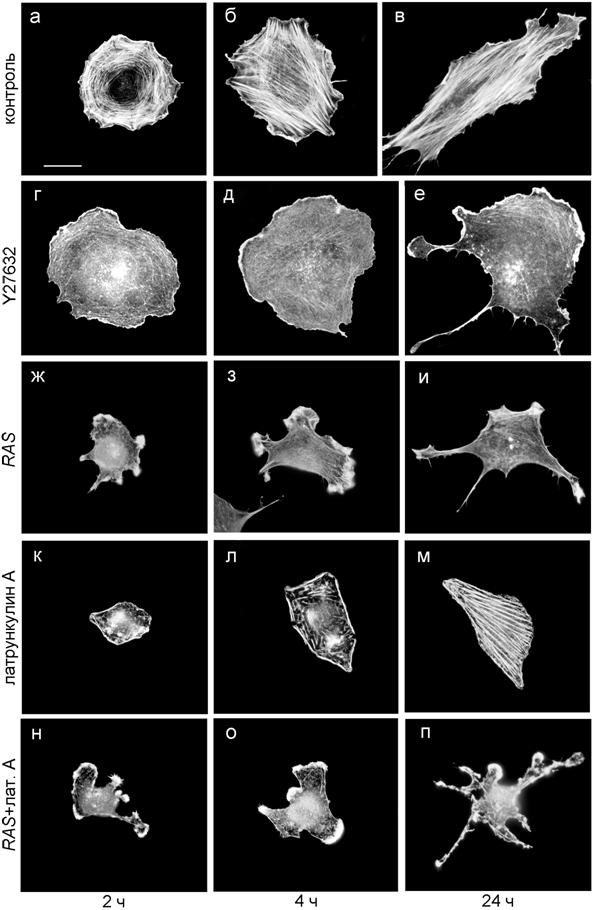 |
Рис. 3. Морфология нормальных и трансформированных клеток на разных этапах распластывания. Флуоресцентная окраска на актин. Масштаб 20 мкм.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
