
Рис. 6. Распределение по расстояниям при миграции в рану нормальных и трансформированных клеток в присутствии латрункулина А и цитохалазина D.
5.3. Эффект ингибиторов полимеризации актина на динамику распластывания.
Действие латрункулина А приводило к серьезным нарушениям распластывания нетрансформированных клеток. На всех сроках края клеток оставались сглаженными, без протрузий. В распластывающихся клетках формировались короткие хаотично расположенные пучки, не образующие единой системы. На всех этапах распластывания площадь обработанных клеток была значительно снижена по сравнению с контролем (табл. 8, рис. 3). Несмотря на подавление распластывания, клетки в присутствии латрункулина могли поляризоваться, и степень их поляризации фактически не менялась.
Табл. 8. Данные морфометрического анализа контуров нормальных клеток через 2, 4 и 24 ч распластывания при обработке ингибиторами полимеризации актина.
площадь, кв. мкм | элонгация | степень поляризации | ||
контроль 2ч | 1551±82 | 0.035±0.005 | 0.256±0.03 | 0.291±0.033 |
4ч | 2541±92 | 0.048±0.007 | 0.521±0.064 | 0.569±0.067 |
24ч | 4665±209 | 0.177±0.022 | 1.178±0.049 | 1.355±0.053 |
лат. A 0.5 мкМ 2ч | 390±17 | 0.015±0.002 | 0.2±0.024 | 0.216±0.024 |
4ч | 769±43 | 0.178±0.031 | 0.745±0.066 | 0.923±0.081 |
24ч | 1498±80 | 0.08±0.015 | 0.887±0.11 | 0.967±0.118 |
цит.D 2ч | 501±25 | 0.074±0.016 | 0.487±0.055 | 0.561±0.059 |
4ч | 813±44 | 0.14±0.019 | 0.578±0.069 | 0.718±0.073 |
24ч | 1990±114 | 0.617±0.09 | 0.997±0.113 | 1.615±0.13 |
При подавлении полимеризации актина в трансформированных клетках не наблюдалось такого снижения скорости распластывания, а конечная площадь снижалась в меньшей степени, нежели площадь нетрансформированных клеток в присутствии ингибиторов (табл. 9).
Эксперименты по отмывке блеббистатина в присутствии латрункулина А показали, что для восстановления/образования как ламеллиподий так и пучков необходима полимеризация актина. Мы полагаем, что в трансформированных клетках баланс интенсивности полимеризации смещен в сторону псевдоподиальной активности за счет ослабления пучков. Возможно, это происходит за счет снижения активности Rho-киназы и повышения активности Rac (Sahai et al, 2001). Поэтому в условиях «дефицита» G-актина под воздействием латрункулина весь свободный актин идет на построение ламеллиподии, тогда как пучки еще больше разбираются. У нормальных же клеток более чувствительным процессом оказывается формирование ламеллиподий, но не пучков.
Табл. 9. Данные морфометрического анализа контуров трансформированных клеток на 2, 4 и 24 ч распластывания при действии латрункулина А и цитохалазина D.
площадь, кв. мкм | дисперсия | элонгация | степень поляризации | |
RAS 2ч | 844±45 | 0,140±0,023 | 0,499±0,057 | 0,639±0,067 |
4ч | 1159±65 | 0,606±0,056 | 0,943±0,087 | 1,548±0,116 |
24ч | 2283±146 | 0,808±0,081 | 1,306±0,114 | 2,115±0,114 |
RAS +лат.A 0.5мкМ 2ч | 692±40 | 0,232±0,063 | 0,648±0,09 | 0,88±0,107 |
4ч | 785±44 | 0,464±0,074 | 0,594±0,106 | 1,057±0,123 |
24ч | 1148±74 | 0,901±0,1481 | 1,046±0,149 | 1,947±0,226 |
RAS +цит. D 2ч | 734±33 | 0,856±0,079 | 1,351±0,152 | 2,208±0,145 |
4ч | 763±37 | 0,5±0,058 | 1,539±0,143 | 2,04±0,12 |
24ч | 1252±68 | 0,897±0,108 | 1,268±0,146 | 2,165±0,163 |
ЗАКЛЮЧЕНИЕ
В нашей работе мы подробно рассмотрели роль актин-миозинового структур в организации движения нормальных и трансформированных клеток. Понижение сократимости является частым характерным признаком клеток, подвергшихся трансформации, и наблюдается, в частности, в случае трансформации фибробластов онкогеном RAS. Мы увидели, что снижение (но не абсолютное подавление) сократимости коррелирует с усилением клеточной подвижности (добавление 45 мкМ блеббистатина, RAS-трансформация) (рис. 7). С другой стороны, мы показали, что минимальная активность миозина необходима для успешного прикрепления клетки к субстрату и эффективной протрузии клеточного края.
Разрушение системы стресс-фибрилл с помощью ингибиторов миозина не оказывает отрицательного влияния на развитие и поддержание поляризации фибробластов на двумерном субстрате. Мы полагаем, что механизм такой поляризации основан на взаимодействии двух сохраняющихся цитоскелетных систем – сети микрофиламентов и МТ. С другой стороны, RAS-трансформированные клетки, занимающие «промежуточное положение» по степени развития стресс-фибрилл между контрольными и обработанными ингибиторами, имеют нарушения поляризации. При высокой общей степени поляризации, они, как правило, мультиполярны: имеют высокую дисперсию, но не элонгацию, и постоянно меняют направление движения.
Мы показали, что нарушения поляризации и направленности движения трансформированных клеток не являются следствием ослабления системы актин-миозиновых пучков (подавление сократимости в нормальных клетках не приводит к нарушению поляризации), но, очевидно, являются следствием нарушения других сигнальных путей, контролируемых онкобелком Ras. Так, снижение сократимости – только один из признаков RAS-трансформированных клеток.
Локомоторное поведение трансформированных клеток оказывается менее чувствительным к действию ингибиторов миозина II и полимеризации актина. Возможно, повышенная активация сигнальных путей у трансформированных клеток за счет постоянной активации Ras, приводит к тому, что они менее эффективно реагируют на внешние сигналы. Такая пониженная реакция трансформированных клеток на регуляторные сигналы, поступающие извне (в нашем случае на воздействие ингибиторов) является существенным адаптационным механизмом, позволяющим опухолевым клеткам инвазировать в соседние ткани, прикрепляться и образовывать новые колонии в условиях, в которых нормальные клетки существовать не могут.
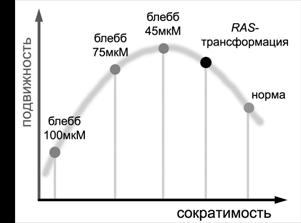 Рис. 7. Схема зависимости клеточной подвижности от сократимости актин-миозинового цитоскелета. При снижении сократимости нормальных клеток с помощью ингибиторов миозина (светлые точки на схеме) клеточная подвижность повышается, но при максимальном ингибировании миозина (добавлении высоких доз ингибитора) нарушается формирование и прикрепление ламеллиподий, что негативно сказывается на подвижности. При RAS-трансформации также снижается сократимость актин-миозина по сравнению с нормой, что усиливает подвижность, кроме того, происходят нарушения регуляции направленности движения, очевидно, не зависящие от изменений сократимости.
Рис. 7. Схема зависимости клеточной подвижности от сократимости актин-миозинового цитоскелета. При снижении сократимости нормальных клеток с помощью ингибиторов миозина (светлые точки на схеме) клеточная подвижность повышается, но при максимальном ингибировании миозина (добавлении высоких доз ингибитора) нарушается формирование и прикрепление ламеллиподий, что негативно сказывается на подвижности. При RAS-трансформации также снижается сократимость актин-миозина по сравнению с нормой, что усиливает подвижность, кроме того, происходят нарушения регуляции направленности движения, очевидно, не зависящие от изменений сократимости.
ВЫВОДЫ
1. При трансформации фибробластов онкогеном RAS актин-миозиновые пучки и фокальные контакты становятся менее выражены. Площадь трансформированных клеток снижается в два раза, они становятся мультиполярны, усиливается подвижность, но клетки двигаются менее направленно.
2. Нетрансформированные фибробласты при ингибировании миозин-зависимой сократимости утрачивают зрелые фокальные контакты и стресс-фибриллы, их подвижность существенно повышается. Однако такие клетки сохраняют способность к развитию поляризации и направленной миграции, что, вероятно, обеспечивается взаимодействием системы микрофиламентов и микротрубочек.
3. Фибробласты, трансформированные онкогеном RAS, имеют общие признаки с клетками, обработанными ингибиторами сократимости. Это нарушение образования стресс-фибрилл и фокальных контактов, усиление псевдоподиальной активности и подвижности, увеличение степени поляризации.
4. В контрольных нетрансформированных клетках миозиновые филаменты имеют высокую степень структурной организации. При RAS-трансформации они становятся менее упорядоченными. При разборке пучков под действием ингибиторов миозина миозиновые филаменты пропадают. Таким образом, в клетке агрегация миозина в биполярные филаменты зависит от того, в какой мере он задействован в стресс-фибриллах, хотя в условиях in vitro миозиновые филаменты могут собираться и без участия актина.
5. В нетрансформированных клетках стресс-фибриллы и фокальные контакты разрушаются при неполном подавлении активности миозина II (45 мкМ блеббистатина). Максимальное подавление активности миозина (100 мкМ бллеббистатина) ведет к ингибированию образования ламеллиподий и фокальных комплексов. Эти процессы взаимозависимы и требуют минимальной активности миозина II. При отмывке блеббистатина первыми восстанавливаются ламеллиподии и фокальные комплексы. Для восстановления и ламеллиподий, и стресс-фибрилл при отмывке необходима полимеризация актина.
6. В нетрансформированных клетках при частичном подавлении полимеризации актина в первую очередь нарушается образование ламеллиподии, а в трансформированных клетках – образование пучков. Таким образом, при RAS-трансформации происходят изменения регуляции полимеризации актина в сторону усиления псевдоподиальной активности и одновременного нарушения построения пучков.
7. Таким образом, онкоген RAS вызывает, во-первых, нарушение регуляции полимеризации актина и снижение актин-миозиновой сократимости, что приводит к усилению подвижности клеток, и, во-вторых, потерю направленности движения, очевидно, обусловленную нарушением других сигнальных путей.
Список работ, опубликованных по теме диссертации
Статьи в журналах, соответствующих Перечню ВАК:
1. Shutova M. S., Alexandrova A. Y., Vasiliev J. M. Regulation of polarity in cells devoid of actin bundle system after treatment with inhibitors of myosin II activity. // Cell Motility and the Cytoskeleton. – 2008. – V.65-9 – P. 734-746.
2. , Александрова исследование распластывания нормальных и трансформированных фибробластов: роль полимеризации микрофиламентов и актин-миозинового сокращения. // Цитология. – 2010. – T.52 №1 – C. 41-51.
Тезисы конференций:
1. Шутова Rho-киназы в морфогенезе и миграции нормальных и трансформированных фибробластов. // Тезисы докладов XII международной конференции студентов, аспирантов и молодых ученых «Ломоносов-2005». –Москва, 2005. – C. 261.
2. Шутова исследование подвижности нормальных и трансформированных фибробластов. Роль актина и миозина 2. // Тезисы докладов XIII международной конференции студентов, аспирантов и молодых ученых «Ломоносов-2006». – Москва, 2006. – C. 254-255.
3. , Александрова ползет клетка. Перестройки цитоскелета, обеспечивающие движение клеток в культуре, роль актина и миозина 2. // Сборник тезисов конференции «Молекулярные механизмы процессов онтогенеза: эмбриогенез, геномы, эволюция» – Москва, 2006. – C. 26.
4. , Шутова роль Arp2/3-зависимой полимеризации актина и формирования первичных адгезионных структур в перестройках актинового цитоскелета, обеспечивающих движение клеток в культуре. // Тезисы конференции «Биология клетки в культуре». – Санкт-Петербург. – Цитология. – 2006. – Т.48 №9 – С. 741.
5. , Александрова подвижности нормальных фибробластов. Роль актин-миозинового сокращения. // Тезисы конференции «Биология клетки в культуре». – Санкт-Петербург. – Цитология. – 2006. – Т.48 №9 – С. 813-814.
6. Shutova M. S., Svitkina T. M. Dynamics of Actin Cytoskeleton Restoration аfter Blebbistatin Treatment. // The American Society for Cell Biology 48th Annual Meeting. – San Francisco, CA, 2008. – Poster # 1125.
7. , , Васильев актинового цитоскелета фибробластов в процессе движения. Нарушения, приводящие к инвазии при опухолевой трансформации. // Материалы ХII Российского онкологического конгресса. – Москва, 2008. – C. 123-124.
8. Shutova M. S., Svitkina T. M. Roles of myosin II in initiation of focal adhesion assembly during their restoration after blebbistatin treatment. // The American Society for Cell Biology 49th Annual Meeting. – San Diego, CA, 2009. – Poster # 1000.
Автор выражает глубокую благодарность Антонине Юрьевне Александровой за активное участие и помощь на всех этапах работы, а также Татьяне Михайловне Свиткиной за предоставленную возможность проведения электронно-микроскопических исследований.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
