

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Неспецифические структуры первого блока по принципу своего действия подразделяются на следующие типы:
♦ восходящие (проводящие возбуждение от периферии к центру);
♦ нисходящие (проводящие возбуждение от центра к периферии).
Восходящие и нисходящие отделы неспецифической системы включают и активационные, и тормозные
пути. В настоящее время установлено, что активационные и тормозные неспецифические механизмы
являются достаточно автономными и независимыми по своей организации на всех уровнях, включая и кору
больших полушарий.
Анатомические особенности неспецифической системы состоят прежде всего в наличии в ней особых
клеток, составляющих ретикулярную (сетчатую) формацию и обладающих, как правило, короткими
аксонами, что объясняет сравнительно медленную скорость распространения возбуждения в этой системе.
Однако в неспецифических структурах обнаружены и длинноаксонные клетки, участвующие в механизме
быстрых активационных процессов.
Корковые структуры первого блока (поясная кора, кора медиальных и базальных, или орбитальных,
отделов лобных долей мозга) принадлежат по своему строению главным образом к коре древнего типа,
состоящей из пяти слоев.
Функциональное значение первого блока в обеспечении психических функций состоит, как уже говорилось
выше, прежде всего в регу-
69
ляции процессов активации, в обеспечении общего активационного фона, на котором осуществляются все психические функции, в поддержании общего тонуса ЦНС, необходимого для любой психической деятельности. Этот аспект работы блока имеет непосредственное отношение к процессам внимания — общего, неизбирательного и селективного, — а также сознания в целом. Внимание и сознание с энергетической точки зрения связаны с определенными уровнями активации. С качественной, содержательной точки зрения они характеризуются набором различных действующих систем и механизмов, обеспечивающих отражение разных аспектов внешнего и внутреннего мира.
Помимо общих неспецифических активационных функций, первый блок мозга непосредственно связан с процессами памяти (в их модально-неспецифической форме), с запечатлением, хранением и переработкой разномодальной информации. Решающее значение этого блока в мнестической деятельности подтверждено многочисленными наблюдениями за больными с поражением срединных неспецифических структур мозга, причем высшие уровни этих структур связаны преимущественно с произвольными формами мнестической деятельности (, 1974а, 1976; Я. К. Киященко и др., 1975; , 1979 и др.). Первый блок мозга является непосредственным мозговым субстратом различных мотивационных и эмоциональных процессов и состояний (наряду с другими мозговыми образованиями). Лимбические структуры мозга, входящие в этот блок (область гиппокампа, поясной извилины, миндалевидного ядра и др.), имеющие тесные связи с орбитальной и медиальной корой лобных и височных долей мозга, являются полифункциональными образованиями. Они участвуют в регуляции различных эмоциональных состояний, прежде всего сравнительно элементарных (базальных) эмоций (страха, удовольствия, гнева и др.), а также мотивационных процессов, связанных с различными потребностями организма. В сложной мозговой организации эмоциональных и мотивационных состояний и процессов лимбические отделы мозга занимают одно из центральных мест. Этот блок мозга воспринимает и перерабатывает разную интероцептивную информацию о состояниях внутренней среды организма и регулирует эти состояния с помощью нейрогуморальных, биохимических механизмов.
Таким образом, первый блок мозга участвует в осуществлении любой психической деятельности, особенно в процессах внимания, памяти, регуляции эмоциональных состояний и сознания в целом.
70
Второй блок — блок приема, переработки и хранения экстероцептивной (т. е. исходящей из внешней среды) информации — включает основные анализаторские системы: зрительную, слуховую и кожно-кинестическую, корковые зоны которых расположены в задних отделах больших полушарий головного мозга. Работа этого блока обеспечивает модально-специфические процессы, а также сложные интегративные формы переработки экстероцептивной информации, необходимой для осуществления высших психических функций. Модально-специфические (или лемнисковые) пути проведения возбуждения имеют иную, чем неспецифические пути, нейронную организацию, им присуща четкая избирательность, проявляющаяся в реагировании лишь на определенный тип раздражителей.
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 43
Все основные анализаторные системы организованы по общему принципу: они состоят из периферического
(рецепторного) и центрального отделов.
Периферические отделы анализаторов осуществляют анализ и дискриминацию стимулов по их физическим
качествам (интенсивности, частоте, длительности и т. п.).
Центральные отделы анализаторов включают несколько уровней, последний из которых — кора больших
полушарий.
Центральные отделы анализируют и синтезируют стимулы не только по физическим параметрам, но и по
сигнальному значению. В целом анализаторы — это аппараты, подготавливающие ответы организма на
внешние раздражители. На каждом из уровней анализаторной системы происходит последовательное
усложнение процессов переработки информации. Максимальной сложности и дробности процессы анализа
и переработки информации достигают в коре больших полушарий. Анализаторные системы
характеризуются иерархическим принципом строения, при этом нейронная организация их уровней
различна.
Кора задних отделов больших полушарий обладает рядом общих черт, позволяющих объединить ее в
единый блок мозга. В ней выделяют «ядерные зоны» анализаторов и «периферию» (по терминологии И. П.
Павлова), или первичные, вторичные и третичные поля (по терминологии ). К ядерным
зонам анализаторов относят первичные и вторичные поля, к периферии — третичные поля. В ядерную зону
зрительного анализатора входят 17, 18 и 19-е поля, в ядерную зону кожно-кинестетического анализатора —
1, 2, 3-е, частично 5-е поля, в ядерную зону звукового анализатора — 41, 42 и 22-е поля,
71
из них первичными полями являются 3, 17 и 41-е, остальные — вторичные (рис. 4, Л).
Первичные поля коры по своей цитоархитектонике принадлежат к коникортикальному, или пылевидному,
типу, который характеризуется широким IV слоем с многочисленными мелкими зерновидными клетками.
Эти клетки принимают и передают пирамидным нейронам III и V слоев импульсы, приходящие по
афферентным проекционным волокнам из подкорковых отделов анализаторов.
Так, первичное 17-е поле коры содержит в IV слое крупные звездчатые клетки, откуда импульсы
переключаются на пирамидные клетки V слоя (клетки Кахала и клетки Майнерта). От пирамидных клеток
первичных полей берут начало нисходящие проекционные волокна, поступающие в соответствующие
двигательные центры местных двигательных рефлексов (например, глазодвигательных). Эта особенность
строения первичных корковых полей (рис. 10) носит название «первичного проекционного нейронного
комплекса коры» (, 1965).
Все первичные корковые поля характеризуются топическим принципом организации («точка в точку»),
согласно которому каждому участку рецепторной поверхности (сетчатки, кожи, кортиевого органа)
соответствует определенный участок в первичной коре, что и дало основание называть ее проекционной.
Величина зоны представительства того или иного рецепторного участка в первичной коре зависит от
функциональной значимости этого участка. Так, область fovea представлена в 17-м поле коры значительно
более широко, чем другие области сетчатки.
Первичная кора организована по принципу вертикальных колонок, объединяющих нейроны с общими
рецептивными полями. Первичные корковые поля непосредственно связаны с соответствующими реле-
ядрами таламуса.
Функции первичной коры состоят в максимально тонком анализе различных физических параметров
стимулов определенной модальности, причем клетки-детекторы первичных полей реагируют на
соответствующий стимул по специфическому типу (не проявляя признаков угасания реакции по мере
повторения стимула).
Вторичные корковые поля по своей цитоархитектонике характеризуются большим развитием клеток,
переключающих афферентные импульсы IV слоя на пирамидные клетки III слоя, откуда берут свое начало
ассоциативные связи коры. Этот тип переключений носит
72
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 44
Рис. 10. Системы связей первичных, вторичных и третичных полей коры
(по Г. И. Полякову):
I — первичные (центральные) поля;
II — вторичные (периферические) поля;
III — третичные поля (зоны перекрытия анализаторов). Сплошной линией выделены системы проекционных (корково-подкорковых) проекционно-ассоциативных и ассоциативных связей коры; пунктиром — другие связи.
1 — рецептор; 2 — эффектор; 3 — нейрон чувствительного узла; 4 — двигательный нейрон; 5, 6 — переключательные нейроны спинного мозга и ствола; 7-10 — переключательные нейроны подкорковых образований; 11, 14 — афферентные
волокна из подкорки; 13 — пирамида V слоя; 16 — пирамида подслоя III; 18 — пирамиды подслоев III2 и III; 12, 15, 17 — звездчатые клетки коры
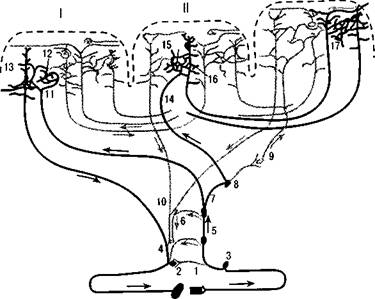
название «вторичного проекционно-ассоциативного нейронного комплекса». Связи вторичных полей коры с подкорковыми структурами более сложны, чем связи первичных полей.
К вторичным полям афферентные импульсы поступают не непосредственно из реле-ядер таламуса, как к первичным, а из ассоциативных ядер таламуса (после их переключения). Иными словами, вторичные поля коры получают более сложную, переработанную информацию с периферии, чем первичные.
73
Вторичные корковые поля функционально объединяют разные анализаторные зоны, осуществляя синтез
раздражений и принимая непосредственное участие в обеспечении различных гностических видов
психической деятельности.
Третичные поля коры задних отделов больших полушарий находятся вне «ядерных зон» анализаторов. К
ним относятся верхнетеменная область (поля 7-е и 40-е), нижнетеменная область (39-е поле), средне-
височная область (21-е и 37-е поля) и зона ТРО — зона перекрытия височной (tempralis), теменной
(parietalis) и затылочной (occipitalis) коры (37-е и частично 39-е поля). Цитоархитектоника этих зон
определяется в известной степени строением соседних ядерных зон анализаторов.
Для третичных полей коры характерен «третичный ассоциативный комплекс», т. е. переключение
импульсов от клеток II слоя к клеткам III слоя (средним и верхним подслоям). Третичные поля не имеют
непосредственной связи с периферией и связаны горизонтальными связями лишь с другими корковыми
зонами.
Третичные поля коры многофункциональны. С их участием осуществляются сложные надмодальностные
виды психической деятельности — символической, речевой, интеллектуальной. Особое значение среди
третичных полей коры задних отделов больших полушарий имеет зона ТРО, обладающая наиболее
сложными интегративными функциями.
Третий блок — блок программирования, регуляции и контроля за протеканием психической деятельности
— включает моторные, премоторные и префронтальные отделы коры лобных долей мозга. Лобные доли
характеризуются большой сложностью строения и множеством двусторонних связей с корковыми и
подкорковыми структурами. К третьему блоку мозга относится конвекситальная лобная кора с ее
корковыми и подкорковыми связями.
Как уже говорилось выше, медиальные и базальные отделы коры лобных долей входят в состав первого —
энергетического — блока мозга. Конвекситальная кора лобных долей мозга занимает 24 % поверхности
больших полушарий. В ней выделяют моторную кору (агранулярную — 4, 6-е поля и слабогранулярную —
8, 44, 45-е поля) и немоторную (гранулярную — 9, 10, 11, 12, 46, 47-е поля). Эти области коры имеют
различные строение и функции. Моторная агранулярная лобная кора составляет ядерную зону
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 45
двигательного анализатора и характеризуется хорошо развитым V слоем, содержащим моторные клетки-пирамиды.
74
Различные участки 4-го первичного поля двигательного анализатора, построенного по соматотопическому принципу, иннервируют разные группы мышц на периферии. В 4-м поле представлена вся мышечная система человека (и поперечно-полосатая, и гладкая мускулатура). Раздражая различные участки 3-го и 4-го полей, У. Пенфилд и Г. Джаспер (1958) уточнили конфигурацию «чувствительного» и «двигательного» человечков — зон проекции и представительства различных мышечных групп (рис. 11, А, Б).
Рис. 11. Схема соматотопической проекции общей чувствительности и двигательных функций в коре головного мозга (по У. Пенфилду):
А — корковая проекция общей чувствительности;
Б — корковая проекция двигательной системы.
Относительные размеры органов отражают ту площадь коры головного мозга,
соответствующие ощущения и движения
с которой могут быть вызваны
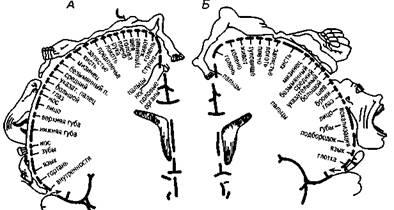
Как видно из рис. 11, Б, «двигательный» человечек имеет непропорционально большие губы, рот, руки, но маленькие туловище и ноги — в соответствии со степенью управляемости теми или иными группами мышц и их общим функциональным значением. «Чувствительный» человечек в целом повторяет строение «двигательного» (рис. 11, А).
В V слое 4-го поля содержатся самые большие клетки ЦНС — моторные клетки Беца, дающие начало пирамидному пути. В 6-м и 8-м полях коры V слой менее широк, но по типу своего строения (наличию пирамид в V и III слоях) эти поля также относятся к моторным агранулярным корковым полям.
75
44-е поле (или «зона Брока») имеет хорошо развитые V и III слои, моторные клетки которых управляют
оральными движениями и движениями речевого аппарата.
Прецентральная моторная и премоторная кора (4, 6, 8-е поля) получает проекции от вентролатеральных ядер
зрительного бугра; префронтальная конвекситальная кора является зоной проекции мелкоклеточной части
ДМ (дорсомедиального) ядра таламуса. В прецентральной (моторной) и премоторной коре берут начало
пирамидный и экстрапирамидный пути. Эти области коры тесно связаны с различными базальными
ганглиями: стриопаллидарной системой, красным ядром, Льюисовым телом и другими подкорковыми
звеньями экстрапирамидной системы.
Префронтальная конвекситальная кора связана многочисленными связями с корой задних отделов больших
полушарий и с симметричными отделами коры лобных долей другого полушария.
Таким образом, многочисленные корково-корковые и корково-подкорковые связи конвекситальной коры
лобных долей мозга обеспечивают возможности, с одной стороны, переработки и интеграции самой
различной афферентации, а с другой — осуществления различного рода регуляторных влияний.
Анатомическое строение третьего блока мозга обусловливает его ведущую роль в программировании
замыслов и целей психической деятельности, в ее регуляции и осуществлении контроля за результатами
отдельных действий, а также всего поведения в целом.
Общая структурно-функциональная модель организации мозга, предложенная , предполагает,
что различные этапы произвольной, опосредованной речью, осознанной психической деятельности
осуществляются с обязательным участием всех трех блоков мозга.
Согласно современным представлениям о психической деятельности, ее структура и процесс протекания
может выглядеть следующим образом:
♦ она начинается с фазы мотивов, намерений, замыслов;
♦ затем эти мотивы, намерения, замыслы превращаются в определенную программу (или «образ результата») действительности, включающую представления о способах ее реализации;
♦ после чего она продолжается в виде фазы реализации этой программы с помощью определенных операций;
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 46
76
Из произведений
Каждый акт поведения — целенаправленное действие, процесс восприятия, запоминания или мышления — опирается на совместную работу этих трех функциональных блоков, причем каждый из них обеспечивает свою сторону нужного процесса. Эти факты стали ясными из длительного систематического анализа тех изменений в психических процессах, которые наступают в результате локальных поражений тех или иных отделов мозга, возникающих в результате травм, опухолей или местных кровоизлияний.
Поражения аппаратов первого блока (стволовых отделов мозга, аппаратов медиальной коры или лимбической области) приводит к модально-неспецифическому снижению тонуса коры и делает избирательное, селективное протекание психической деятельности невозможным или очень трудным.
Поражение аппаратов второго блока (вторичных отделов левой височной или теменно-затылочной области) существенно нарушает условия, необходимые для приема и переработки информации, причем каждое из этих поражений приводит к отчетливым модально-специфическим (зрительным, слуховым, пространственно-кинестетическим) нарушениям, а поражения этих аппаратов левого полушария коренным образом ограничивают возможность переработки соответствующей информации при помощи языка, ==>
♦ завершается психическая деятельность фазой сличения полученных результатов с исходным «образом результата». В случае несоответствия этих данных психическая деятельность продолжается до получения нужного результата.
Эта схема (или психологическая структура) психической деятельности, многократно описанная в трудах (1972) и других отечественных и зарубежных психологов (, 1967; К. Прибрам, 1975 и др.), в соответствии с моделью «трех блоков» может быть соотнесена с мозгом следующим образом.
1. В начальной стадии формирования мотивов в любой сознательной психической деятельности (гностической, мнестической, интеллектуальной) принимает участие преимущественно первый блок мозга. Он обеспечивает также оптимальный общий уровень активности мозга и осуществление избирательных, селективных форм активности, необходимых для протекания конкретных видов психической деятельности. Первый блок мозга преимущественно ответствен и за эмоциональное «подкрепление» психической деятельности (переживание успеха-неуспеха).
77
Из произведений
► Наконец, поражение аппаратов третьего блока (лобных отделов мозга), не изменяя общего тонуса коры и не затрагивая
основных условий процесса приема информации, существенно нарушает процесс ее активной переработки, затрудняя
процесс возникновения намерений, программирования действий, препятствует стойкой регуляции и контролю за их
протеканием.
Легко видеть, что каждая из этих форм поражений мозга устраняет то или иное условие, необходимое для нормального
протекания сознательной деятельности, и приводит к ее дезинтеграции, к распаду организованно работающих
функциональных мозговых систем.
Едва ли наиболее существенным является, однако, тот факт, что каждое из таких поражений нарушает сознательную
деятельность особым, специфическим образом, так что распады функциональных систем, возникающие в результате
снижения тонуса коры, в результате нарушения модально-специфических форм обработки информации или в результате
нарушения регулярной деятельности, резко отличаются друг от друга.
(. Основные проблемы нейролингвистики. — М.: МГУ, 1975. - С. 47-48.)
2. Стадия формирования целей, программ деятельности связана преимущественно с работой третьего блока мозга, так же как и стадия контроля за реализацией программы.
3. Операциональная стадия деятельности реализуется преимущественно с помощью второго блока мозга. Поражение одного из трех блоков (или его отдела) отражается на любой психической деятельности, так как приводит к нарушению соответствующей стадии (фазы, этапа) ее реализации.
Данная общая схема функционирования мозга как субстрата сложных сознательных форм психической деятельности находит конкретное подтверждение при нейропсихологическом анализе нарушений высших психических функций, возникающих вследствие локальных поражений головного мозга.
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 47
Глава 4. Проблема межполушарной асимметрии мозга и межполушарного взаимодействия
Проблема межполушарной асимметрии и межполушарного взаимодействия — одна из наиболее актуальных в современном естествознании. В настоящее время она разрабатывается разными нейронауками: нейроанатомией, нейрофизиологией, нейробиологией и др. Весьма продуктивно она изучается и нейропсихологией. Локальные поражения мозга в качестве основной модели для нейропсихологических исследований открывают уникальные возможности изучения данной проблемы на человеке. Все возрастающее число публикаций, посвященных оценке функций левого и правого полушарий мозга у человека, их роли в различных видах психической деятельности, свидетельствует о широком научном интересе к этому направлению исследований.
Межполушарная асимметрия представляет собой одну из фундаментальных закономерностей работы мозга не только человека, но и животных (, 1975, 1989; , 1979; С. Спрингер, Г. Дейч, 1983 и мн. др.). Однако, несмотря на сравнительно длительную историю изучения данной проблемы (ее начало можно отнести к 1861 году, когда П. Брока открыл «центр» речевой моторики в левом полушарии головного мозга) и огромное количество современных публикаций по различным ее аспектам (биологическим, морфологическим, физиологическим, экспериментально-психологическим, клиническим, лингвистическим и др.), сколько-нибудь законченной теории, объясняющей функциональную асимметрию больших полушарий и учитывающей действие как генетических, так и социокультурных факторов в ее формировании, пока не существует.
Фактические данные, полученные на разном клиническом и экспериментальном материале, многочисленны и нередко противоречивы. Можно сказать, что накопление фактического материала по данной проблеме явно опережает его теоретическое осмысление.
79
В настоящее время получено множество фактов о неравнозначности левого и правого полушарий головного мозга по различным показателям. Это и анатомические, и физиологические данные, и материалы наблюдений за больными, имеющими сходные поражения левого и правого полушарий. Анатомические данные, полученные Московским Институтом Мозга РАМН, а также результаты зарубежных авторов свидетельствуют, что уже у животных (крыс, кошек, обезьян и др.) имеются анатомические различия в строении левого и правого полушарий мозга. Наиболее отчетливы они в височной области. Так, было показано, что у шимпанзе и макаки-резуса длина Сильвиевой борозды в левом полушарии больше, чем в правом (, 1976, 1983 и др.).
В Институте Мозга РАМН специально изучался вопрос о «структурных предпосылках» межполушарной асимметрии мозга человека (в соответствии с терминологией, введенной ). Еще в 30-40-е годы XX века сотрудники этого института (, , и др.) обнаружили следующие структурные различия полей правого и левого полушарий:
♦ общая площадь нижней лобной извилины (45-е поле) у правшей слева больше, чем справа;
♦ в нижнетеменных областях коры (39-е и 40-е поля) слева увеличены размеры коры в глубине борозд;
♦ островковая область слева больше, чем справа;
♦ задняя оперкулярная зона (или зона Вернике) в височной области в левом полушарии на одну треть больше, чем в правом (рис. 12, А, Б);
♦ отмечается морфологическая асимметрия сосудов средней мозговой артерии в левом и правом полушариях;
♦ длина левого полушария превышает длину правого более чем в 54 % случаев (по данным , 1981);
♦ степень вертикальной упорядоченности поперечника коры, прежде всего III слоя (богатого ассоциативными связями), достоверно выше в корковых полях мозга человека по сравнению с высшими приматами и существенно выше в нижнелобных (44-е, 45-е поля) и височных (22-е, 41-е поля) областях левого полушария по сравнению с правым.
Изучение структуры полей коры у человека на нейронном уровне также выявило латеральные различия. Установлено, что:
80
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 48
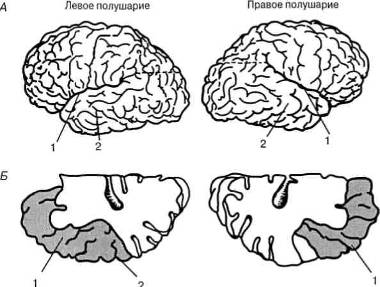
Рис. 12. Анатомическая асимметрия коры мозга человека:
А — Сильвиева борозда, которая определяет верхнюю границу височной доли (1), поднимается
более круто в правом полушарии по сравнению с левым; 2 — височная область;
Б — верхняя часть височной доли (1 ) обычно значительно больше в левом полушарии по сравнению с правым. Эта
область в левом полушарии составляет часть зоны Вернике (2), играющей важную роль в мозговой организации речевых
процессов (по Н. Гешвинду)
♦ размеры нейронов III и IV слоев в 44-м и 45-м полях в левом полушарии больше, чем в правом;
♦ размеры гигантских пирамидных клеток Беца в V слое 4-го моторного поля в левом полушарии также превышают размеры этих нейронов в правом полушарии (R. С. Truex, M. В. Carhenter, 1964; «Asymmetrical...», 1978 и др.).
Имеются данные о морфологическом различии в организации левого и правого таламусов, а также левого и правого хвостатых ядер. Особенно четкая асимметрия строения наблюдается в ядрах таламуса, связанных с речевыми функциями (например, в заднем латеральном ядре, которое имеет проекции к задней височно-теменной и к нижнетеменной коре).
Таким образом, большинство исследователей убеждены в существовании морфологической основы функциональной асимметрии мозга,
81
которая является структурным основанием функциональных различий.
Межполушарная асимметрия мозга является также объектом многочисленных физиологических исследований, которые проводятся как у нас в стране, так и за рубежом.
Многими авторами изучаются ЭЭГ-проявления функциональной межполушарной асимметрии мозга как в состоянии покоя, так и во время психической деятельности. При изучении межполушарных различий ЭЭГ в состоянии покоя одни исследователи отмечают большую выраженность депрессии альфа-ритма в левом полушарии по сравнению с правым, другие считают альфа-компоненты спектра ЭЭГ относительно симметричными. Однако, по данным многих авторов, при интеллектуальной деятельности межполушарная асимметрия по показателям альфа-ритма усиливается. Большинство авторов утверждают, что во время интеллектуального напряжения альфа-ритм по амплитуде, индексу или суммарной энергии выражен в левом полушарии слабее, чем в правом. и (1977) выявили правостороннее доминирование альфа-депрессии при наглядно-образных формах деятельности и левостороннее — при вербальных. По данным и его сотрудников (, 1980 и др.), у здоровых испытуемых наблюдаются различные паттерны межполушарной асимметрии (по показателям альфа - и бета-ритмов) при выполнении разных видов деятельности. При переходе от невербальных заданий к вербальным происходит уменьшение правополушарного доминирования реакции активации или смена правополушарного доминирования на левополушарное. Усиление межполушарной асимметрии биопотенциалов и связь паттернов активации с характером интеллектуальной деятельности были выявлены и при использовании показателей пространственной синхронизации биопотенциалов («Проблемы нейропсихологии», 1977 и др.).
Важное место в физиологических исследованиях проблемы функциональной асимметрии полушарий занимает метод регистрации вызванных потенциалов (ВП). По данным (1983, 1993), ВП в задних отделах правого полушария опережают по времени ВП в тех же отделах левого полушария. Это расценивается как доказательство осуществления в правом полушарии первичного зрительно-пространственного анализа стимулов. Уровень межполушарной асимметрии ВП (при прочих равных условиях) зависит от характера стимула и области регистрации ответа: при предъявлении как вербальных,
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 49
так
82
и пространственно-структурных стимулов асимметрия ВП максимально выражена в височных областях коры. Асимметрия зрительных ВП на сложные стимулы (предметные изображения, наложенные друг на друга) обнаружена в теменно-затылочных и в премоторных областях мозга, причем при трудных для испытуемого заданиях степень асимметрии ВП возрастает («Проблемы нейропсихологии», 1977 и др.). Асимметрия биоэлектрической активности левого и правого полушарий мозга в настоящее время изучается в разных научных центрах с использованием следующих новых современных методов:
♦ метода измерения локального мозгового кровотока;
♦ томографических методов (позитронно-эмиссионной, магнитно-резонансной томографии и др.);
♦ метода термоэнцефалоскопии и ряда других (см. H. H. Данилова, 1998).
Полученные современной психофизиологией данные свидетельствуют о том, что лево-правая асимметрия
биопотенциалов характерна для нормы и проявляется особенно четко в условиях психической деятельности
(рис. 13; цветная вклейка).
Асимметрия биопотенциалов является региональным свойством и зависит от характера выполняемой
деятельности. Безусловна связь типа и степени асимметрии биопотенциалов с индивидуальным «профилем»
латеральной организации мозга испытуемого (правшеством-левшеством).
Клинические наблюдения за больными с локальными поражениями левого и правого полушарий мозга дают
богатый фактический материал о функциональной неравноценности полушарий. Начиная с открытия П.
Брока моторного «центра речи» в левой нижнелобной области и до настоящего времени клиника локальных
поражений головного мозга предоставляет все новые разнообразные доказательства функциональной
асимметрии полушарий. К ним относятся прежде всего следующие:
♦ многочисленные данные о появлении речевых нарушений (афазий) при поражении левого полушария (преимущественно у правшей);
♦ факты о ведущей роли левого полушария в осуществлении не только речевых, но и других связанных с речью функций.
Результаты этих исследований обобщены в работах (1962, 1973 и др.), в которых на большом клиническом материале обосновывается функциональное значение различных отделов левого по-
83
лушария головного мозга в организации речи и других высших психических функций. Специальному анализу подвергались клинические материалы о связи между доминантностью полушария по речи и ведущей рукой. Выяснилось, что далеко не во всех случаях эти функции совпадают и что возникновение афазии при поражении левого полушария наблюдается не только у правшей, но и у некоторых левшей и амбидекстров.
Клинические наблюдения специфики нарушений психических функций при локальных поражениях левого и правого полушарий в последние годы подкрепляются и специальными исследованиями с использованием следующего инструментария:
♦ хирургических методов, направленных на «расщепление мозга» (рис. 14, А, Б);
♦ метода односторонней электрошоковой терапии;
♦ метода Вада (введение амитала натрия в одну из сонных артерий).
Эти методы открывают новые возможности для изучения функций левого и правого полушарий мозга и
роли объединяющих их комиссур (мозолистого тела и др.).
Таким образом, в настоящее время собран огромный эмпирический материал, подтвержденный данными
анатомии, физиологии, а также клиническими исследованиями, свидетельствующий о неравнозначности
структур и функций левого и правого полушарий головного мозга человека.
Развитие теории межполушарной асимметрии мозга происходило в несколько этапов.
На первом этапе многие ученые считали, что левое полушарие является полностью доминантным по
отношению к речи, мануальным функциям, а также другим высшим психическим процессам. Правому
полушарию отводилась второстепенная, подчиненная роль в реализации всех психических процессов (J. G.
Beaumont, 1974; «Lateralization...», 1977; E. A. Zillmer et al., 2001 и др.). Концепция левополушарной
доминантности была основана на положении об абсолютной противоположности функций левого и
правого полушарий мозга; при этом сама доминантность понималась как исключительная роль левого
полушария в обеспечении речи и других высших связанных с ней психических функций.
Накопление фактов относительно участия правого полушария головного мозга в речевой деятельности, а
левого — в невербальных,
84
Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 50
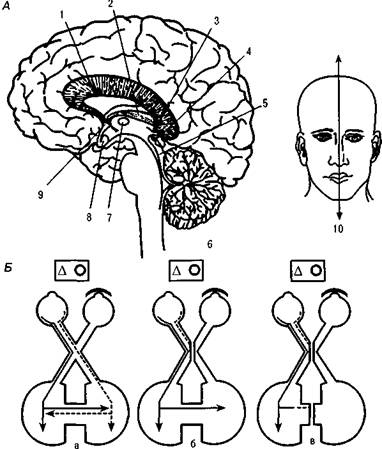
Рис. 14. Межполушарные связи:
А — основные межполушарные комиссуры:
1 — мозолистое тело,
2 — гиппокампальная комиссура, 3 — уздечка, 4 — задняя комиссура, 5 — связи четверохолмия, 6 — мозжечок, 7 — промежуточная масса, 8 — зрительная хиазма, 9 — передняя комиссура, 10 — линия рассечения комиссур мозга (по Р. У. Сперри);
Б — поступление зрительной информации от каждого глаза в оба полушария в норме (а), нарушение взаимодействия полушарий после рассечения хиазмы (б); прекращение взаимодействия полушарий после рассечения и других комиссур (в) (по Д. Брэдшоу и Н. Нетлетону)
85
Из произведений
...вопрос о совместной работе левого и правого полушарий в осуществлении сложных форм психической деятельности человека является в настоящее время едва ли не наиболее дискутируемым в нейропсихологии.
Начиная с известного открытия Брока считалось, что левое (доминантное у правшей) полушарие связано с речью и оно обеспечивает протекание сложных форм психической деятельности человека, в которых речь играет решающую роль. Функция правого (субдоминантного у правшей) полушария оставалась неясной, и лишь отдельные разнообразные факты указывали на его тесную связь с осуществлением не связанных с речью процессов, в первую очередь с мозговой организацией процессов восприятия.
Однако за последние десятилетия накопилось большое число клинических и психологических фактов, которые заставляют пересмотреть это, в основном верное, положение.
Применяя значительно более точные методы (к которым относятся введения амитала натрия в левую и правую сонные артерии, позволяющее на краткий срок избирательно исключать из работы левое или правое полушарие, метод изучения дихотического слуха, дающий возможность точно оценивать доминантность полушарий, и др.), удалось установить, что доминантность левого полушария у правшей вовсе не столь абсолютна, как это считалось раньше. Было показано, что существует парциальная доминантность левого полушария и что люди, у которых левое полушарие доминантно по функциям речи, могут проявлять признаки доминантности правого полушария по другим показателям. Было обнаружено также, что диапазон людей, занимающих промежуточное место по степени доминантности левого полушария, значительно шире, чем это предполагалось, и что ==>

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 |
