

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Отобразив данные из табл. 2 и 3 на координатной плоскости, где по оси абсцисс отложены значения времени в млн лет назад, а по оси ординат отложены значения ИЭ, и объединив полученные области значений, автор получил график изменения во времени индекса энцефализации таксона «прямоходящие» (рис. 1). Названия видов и условные обозначения зоологических объектов, соответствующие рассчитанным областям значений и отдельным величинам ИЭ, приведены на рис. 1 в виде сносок. Координаты и значение точки «HOMO» будут разъяснены в гл. V.
Таблица 3
Область значений индекса энцефализации для современных
человекообразных обезьян
№ | Вид обезьян | Вес тела, кг | Объём мозговой полости, см3 | Индекс энцефализации |
1 | Шимпанзе | 50 | 400 | 2.7 |
2 | Орангутанг | 150 | 400 | 0.9 |
Описание полученного графика (см. рис. 1). Предварительное рассмотрение графика показывает, что он состоит из трёх областей значений:
1) область значений «I» построена для современных понгид в предположении, что для них и для их миоценовых экологических эквивалентов – той части гоминоидов, из которой предположительно возникли австралопитеки, величина ИЭ будет примерно одинакова;
2) область значений «IIIa, b,c» представляет изменение значения ИЭ во времени для собственно таксона «прямоходящие». Общее увеличение ИЭ области значений «III» относительно области значений «I» объясняется, вероятно, увеличением мозга, связанным с переходом от брахиации или получетвероногой наземной локомоции к бипедии, и соответственно с переходом в другую эколого-энергетическую нишу;
3) область значений «II» построена простым соединением областей «I» и «III» в интервале времени 8–7 млн лет, отражающем переход к прямохождению в соответствии с работой [Вырский, 1989];
Анализ области значений ИЭ таксона «прямоходящие». Область значений ИЭ для таксона «прямоходящие», обозначенная на графике (см. рис. 1) как область «IIIa, b,c», показывает значительную неоднородность и позволяет условно разделить её на три меньшие области значений.
Хронологическое рассмотрение показывает, что с момента определяемого наиболее ранним проявлением таксона (6.2–5.65 млн лет), функция ИЭ проходит параллельно оси абсцисс не изменяясь до 3 ‑ 2.5 млн лет. Эту область значений индекса обозначим как область «IIIa».
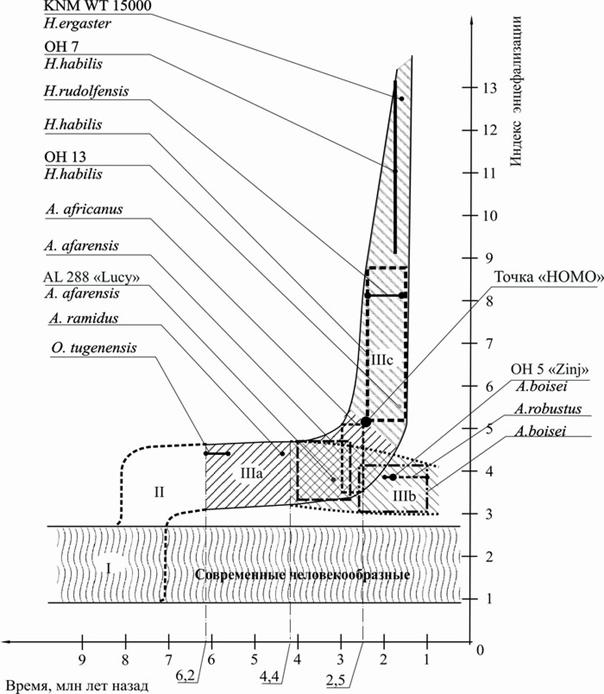
Рис. 1. График изменения во времени индекса энцефализации
таксона «прямоходящие»
Начиная с 3–2.5 млн лет наблюдается заметное разделение графика на две области значений: область «IIIb», в которой значение ИЭ не меняется или даже несколько уменьшается, и область «IIIc», характеризуемую значительным, практически экспоненциальным увеличением значения индекса.
Ввиду того, что признак «индекс энцефализации» не позволяет удовлетворительно разделить таксон «прямоходящие» в соответствии с принципом, указанным во введении, на два симпатрических, необходимо предварительно разделить его на основании других определительных признаков.
IV. расчленение таксона «прямоходящие»
по специализации в диете
Признак «растительноядная специализация». Область значений «IIIb», выделенная на графике (см. рис. 1), представлена двумя видами – A. robustus и A. boisei, входящими в таксон «прямоходящие». Обращаясь к работе [Вырский, 1989], отметим, что эти виды характеризуются адаптацией к растительноядности, которая выражается дробящей и перетирающей специализацией зубочелюстного аппарата.
На костных остатках это определяется своеобразным соотношением переднего и заднего отделов челюсти: мелкие резцы, небольшой клык, функционально близкий к премолярам, моляризованные нижние премоляры, очень крупные моляры. Кроме того, для мегагнатных форм растительноядных характерно особое строение мыщелков, позволяющее производить жевательные движения нижней челюстью в горизонтальной плоскости. Растительноядность сопровождается наличием саггитального гребня, служащего для крепления мощных боковых жевательных мышц.
Если растительноядную специализацию принять за интегрированный определительный признак, то эти два вида – A. robustus и A. boisei – можно объединить в таксон более низкого уровня с названием «прямоходящие растительноядные», отражающим и таксоноопределяющий признак, и вхождение в состав таксона «прямоходящие».
Обращаясь к видам, расположенным в области значений «IIIa» графика (см. рис. 1), мы также обнаруживаем образцы, удовлетворяющие этому признаку. Так, отдельные образцы вида A. ramidus возрастом 4.4 млн лет показывают редукцию передних зубов, износ клыков в окклюзивной плоскости и, наряду с образцом KNM-KP 29281 (нижняя челюсть с зубным рядом) вида A. anamensis возрастом 4.2–3.9 млн лет, представляют наиболее раннее проявление сформировавшейся растительноядной специализации. Признакам растительноядной специализации удовлетворяет также образец BOU-VP-17/1 (нижняя челюсть с зубным рядом) вида A. garhi возрастом 2.5 млн лет.
Не проводя ревизии и идентификации по специализации к растительноядности всех образцов таксона «прямоходящие», включим в состав таксона «прямоходящие растительноядные» оригинальный вид A. ramidus (4.4 млн лет), образец KNM-KP 29281 вида A. anamensis (4.2‑3.9 млн лет), образец BOU-VP-17/1 вида A. garhi (2.5 млн лет) и два вида – A. robustus и A. boisei (общим возрастом 2.5–1.2 млн лет).
Заметим, что члены этого таксона расположены во времени диахронно, последовательно, представляя, таким образом, единую эволюционную линию или так называемую «эволюционную непрерывность», что в свою очередь свидетельствует о том, что зоологическим объектам, входящим в таксон «прямоходящие растительноядные», свойственно генетическое единство – один из триады признаков вида, биологической концепции вида.
Признак «всеядная специализация». Возвращаясь к области «IIIa», выделенной на графике (см. рис. 1), обращаем внимание на то, что образцы, имеющие хорошую сохранность зубочелюстного аппарата, если только они не показывают адаптации к растительноядности, уверенно идентифицируются с всеядной специализацией.
Всеядность проявляется на костных остатках в виде альвеолярного прогнатизма, обеспечивающего «скусывающую» функцию – псалидонтность. Соответственно этому обнаруживается наличие лингвального износа верхних резцов и клыков и лабиального износа нижних. Увеличенный клык, выступающий из зубного ряда, наличие диастем, увеличенный надглазничный валик – всё это свидетельствуют о значительной доли мясной пищи в рационе. Саггитальный гребень у «всеядных» отсутствует.
Повторяем, что в рамках данной работы автор не ставил задачи полной ревизии ископаемого материала для идентификации по специализации в диете, покажем лишь, что верхний резец (образец Bar-1001'00) и клык (BAR-1425'00) вида Orrorin tugenensis возрастом 6.2–5.65 млн лет представляют наиболее раннее проявление сформировавшейся всеядной специализации. Размер и форма передних зубов, наличие диастем и другие признаки верхней челюсти (образец AL 200-1a) вида A. afarensis возрастом 3 млн лет также показывают всеядность.
Анализ строения зубочелюстного аппарата оригинальных видов рода Homo из области значений «IIIc» также показывает полное их тождество с образцами специализированными к всеядности, что позволяет внутри таксона «прямоходящие» выделить второй аллохронный таксон с названием «прямоходящие всеядные», также отражающим и таксоноопределяющий признак, и вхождение в состав таксона более высокого уровня.
Таким образом, в состав таксона «прямоходящие всеядные» включены образцы Bar-1001'00 и BAR-1425'00 вида Orrorin tugenensis (6.2–5.65 млн лет), образец AL 200-1a вида A. afarensis (3 млн лет) и три вида рода Homo – H. rudolfensis (2.4–1.8 млн лет), H. habilis (2.3–1.5 млн лет) и H. ergaster (1.8–1.2 млн лет).
Распределение во времени членов таксона показывает диахронное, последовательное во времени расположение объединяемых образцов и видов, которые представляют единую эволюционную линию или так называемую «эволюционную непрерывность», и поэтому таксону «прямоходящие всеядные», так же как и таксону «прямоходящие растительноядные», свойственно генетическое единство – один из триады признаков вида, биологической концепции вида.
V. Систематика прямоходящих
периода 6.2–1.2 млн лет назад
1. Все африканские прямоходящие периода 6.2–1.2 млн лет назад, объединённые для целей настоящего исследования в аллохронный таксон «прямоходящие» (см. табл. 1), разделены по специализации в диете на два таксона более низкого уровня – «прямоходящие растительноядные» возрастом 4.4–1.2 млн лет и «прямоходящие всеядные» возрастом 6.2 ‑ 1.2 млн лет, каждый из которых представляет независимую эволюционную, филетическую линию, удовлетворяющую триаде признаков [Майр, 1971], определяющих вид в биологической концепции вида:
во-первых, они занимают различные эколого-энергетические ниши;
во-вторых, в условиях симпатрического сосуществования на одном континенте в течение достаточно длительного промежутка времени, по крайней мере от 4.4 до 1.2 млн лет назад, популяции этих таксонов, не смешиваясь, эволюционировали каждая в свою сторону, что говорит о наличии генетической изоляции их друг от друга, или о репродуктивном сообществе внутри популяций каждого из этих двух таксонов;
в-третьих, и это уже было сказано выше, каждый из этих таксонов имеет собственный независимый генофонд, представляя генетическое единство.
Таким образом, таксон «прямоходящие» состоит из двух таксонов видового уровня, специализирующихся в различных системах питания. Для удобства дальнейшего изложения присвоим этим видам рабочие названия, соответствующие названиям таксонов, из которых они состоят: «прямоходящие растительноядные» и «прямоходящие всеядные» соответственно.
Ввиду того, что на начало исследуемого периода (6.2 млн лет) признак «прямохождение» проявился полностью сформированным, можно считать, что его возникновение произошло в более раннее время.
То же самое можно сказать и о признаке «всеядность», поскольку и он на начало исследуемого периода (6.2 млн лет) проявился полностью сформированным.
Признак «растительноядность» проявился на ископаемых образцах, начиная с 4.4 млн лет, и формально допустимо ответвление растительноядных от всеядных в интервале времени от 6.2 до 4.4 млн лет, однако полное отсутствие каких-либо переходных форм в этот период времени не опровергает и модели, в которой возраст 4.4 млн лет примерно соответствует времени прихода «растительноядных» из Азии на африканский континент [Вырский, 1989].
В соответствии с этими представлениями следует вывод, что таксономическо-систематическое понятие «прямоходящие» охватывает филетический ряд, начинающийся от ещё неизвестного вида-основателя из надсемейства гоминоидов, перешедшего к прямохождению, двух видов –«прямоходящие растительноядные» и «прямоходящие всеядные», на которые он впоследствии разделился и которые уже сформированными представлены на африканском континенте периода 6.2–1.2 млн лет под общим названием «австралопитеки».
2. Возвращаясь к таксону/виду «прямоходящие всеядные», представленному на графике (см. рис. 1) в виде диахронной последовательности двух областей значений «IIIa» и «IIIc», мы должны обратить внимание на резкий характер изменения ИЭ, произошедший 3–2.5 млн лет назад. Перегиб функции, представляя начало экспоненциального увеличении ИЭ и соответственно начало относительного увеличения объёма головного мозга, совпадает по времени с появлением на африканском континенте первых свидетельств трудовой деятельности – каменной индустрии Хадар [Джохансон, Иди, 1984].
Если отказаться от необходимости симпатрического деления таксона, выраженного в заявленном принципе подбора таксоноопределяющих признаков, то можно применить граничное значение величины ИЭ в точке перегиба как признак, разделяющий таксон/вид «прямоходящие всеядные» на два диахронных таксона, принадлежащих единой эволюционной (вертикальной) непрерывности.
Определим это граничное значение ИЭ на графике (см. рис. 1) условной точкой ~5.15 по оси ординат, что соответствует значению ~2.5 млн лет по оси абсцисс, разделяющей верхнее значение области ИЭ для оригинального вида A. africanus (5.1) и нижнее значение области ИЭ оригинального вида H. habilis (5.2).
Вновь образованным таксонам присвоим названия, соответствующие названиям областей на графике (см. рис. 1), – таксон «IIIa» и таксон «IIIc». В таксон «IIIa», характеризуемый признаком «величина ИЭ менее 5.15», включим образцы Bar-1001'00 и BAR-1425'00 вида Orrorin tugenensis (6.2‑5.65 млн лет) и AL 200-1a вида A. afarensis (3 млн лет). В таксон «IIIc», характеризуемый признаком «величина ИЭ более 5.15», включим оригинальные виды H. rudolfensis, H. habilis и H. ergaster.
Рассмотрение таксона «IIIc» с точки зрения биологической концепции вида показывает наличие у него триады признаков вида:
во-первых, налицо переход таксона «IIIc» в новую эколого-энергетическую нишу, связанную с применением изготовленных орудий труда;
во-вторых, такого рода нарушение онтогенетической стабильности, выраженное в изменении как самого головного мозга, так формы и размеров краниальных костей, заметно отличающихся от таковых материнского таксона/вида «IIIa», безусловно, имеет генотипический характер, так что можно говорить о некоей генетической изоляции таксона «IIIc» от материнского таксона/вида «IIIa», формально, правда, не доказуемое из-за отсутствия образцов «всеядных» моложе 2.5 млн лет со значением ИЭ менее 5.15;
в-третьих, экспоненциальное изменение ИЭ таксона «IIIc» на графике (см. рис. 1) отражает уникальное дивергентное видообразование, позволяющее говорить о независимом генофонде и генетическом единстве новой эволюционной ветви.
Таким образом, с точки зрения биологической концепции вида мы наблюдаем дивергентное возникновение нового вида в рамках одной филетической линии (эволюционной непрерывности). В состав нового вида включаем оригинальные виды: H. rudolfensis, H. habilis и H. ergaster, входящие в состав таксона «IIIc». Видоопределяющим признаком нового вида можно считать интегральную триаду признаков: два систематических признака, присущих материнскому виду, – прямохождение и всеядность и третий, приобретённый признак, – относительное увеличение размеров головного мозга, характеризуемое значением ИЭ выше 5.15. Виду присвоить рабочее название «IIIc».
Учитывая эволюционный переход в новую эколого-энергетическую нишу, определяемую применением изготовленных орудий труда, а также предположительное дальнейшее разветвление по крайней мере на два биологических вида, используя рекомендацию Э. Майра [Майр, 1971], а также с целью сохранения систематической функции вертикальной палеонтологической классификации приматов, автор считает необходимым назначить новый род, основателем которого и в состав которого в исследуемом периоде входит вид с рабочим названием «IIIc».
Завершая рассмотрение систематики прямоходящих африканского континента возрастом 6.2–1.2 млн лет, можно констатировать существование двух независимых эволюционных, филетических линий соответственно двух таксонов видового уровня – «прямоходящие растительноядные» возрастом 4.4–1.2 млн лет и «прямоходящие всеядные» возрастом 6.2 ‑ 1.2 млн лет.
В момент времени ~2.5 млн лет наблюдается дивергентное преобразование таксона/вида «прямоходящие всеядные» и возникновение нового таксона видового уровня (с точки зрения биологической концепции вида) и родового уровня (с точки зрения вертикальной палеонтологической классификации), идентифицируемые относительным увеличением размеров головного мозга и характеризуемые появлением первичного социального признака – трудовая деятельность.
Для удобства дальнейших исследований и отражения уникальности произошедшего видообразования точку на оси абсцисс 2.5 млн лет графика (см. рис. 1) со значением ИЭ 5.15 на оси ординат следует назвать точкой «HOMO».
VI. Классификация африканских прямоходящих
приматов периода 6.2–1.2 млн лет
Для удобства дальнейших исследований и упорядочения классификации автор считает необходимым отразить результаты, полученные на основании данного исследования, в номенклатуре рассмотренных выше оригинальных видов.
1. Род «Прямоходящие»
Таксоноопределяющему интегрированному признаку «прямохождение» присвоить ранг родоопределяющего признака для рода с названием «Bipedius», куда включить образцы 14 оригинальных видов, объединённых в таксон «прямоходящие» и показанных в табл. 1.
Вновь образованный род включить в зоологическую номенклатуру отряда приматов следующим образом:
Палеонтологическая систематика:
· Отряд Primates Linnaeus, 1758
- Подотряд Anthropoidea Mivart, 1864 Надсемейство Hominoidea Gray, 1825 Bipedius, genus novum
Этимология
Слово Bipedius в переводе с латыни означает двуногий и соответствует как способу передвижения, при котором передние конечности освобождены от функции опоры при стоянии и передвижении, так и родообразующему признаку, необходимому и достаточному для включения примата надсемейства гоминоидов в этот род.
Описание рода Bipedius
Род двуногих приматов характеризуется морфологией, определяемой прямохождением и неподвижным стоянием на двух (нижних конечностях) ногах, при котором плечевой комплекс и соответственно грудные конечности свободны от функции локомоции или опоры. Во внешнем виде это проявляется, прежде всего, в выпрямленном положении тела, вертикальности туловища, при полной разогнутости тазобедренного сустава и нижних конечностей в коленном сочленении [Нестурх, 1970].
Типовым видом, связывающим установленный род Bipedius с зоологическими объектами, назначается оригинальный вид A. afarensis, представленный образцом AL 288-1, найденным D. Johanson и T. Gray в 1974 г. в Хадаре Эфиопии и названным «Lucy». Образец наиболее полно скомплектован, наиболее полно отражает прямохождение исследуемого периода, являясь, таким образом, носителем родообразующего признака, отличающего род от протоморфного, материнского вида.
Ареал рода
Африканский континент
Горизонты залегания
Возраст слоёв залегания определён во временном интервале 6.2 ‑ 1.2 млн лет. Однако учитывая, что в этом временном интервале род проявился на африканском континенте не только сформированным, но и уже разделившимся на два биологических вида, а также учитывая результаты исследования [Вырский, 1989], в геохронологический интервал рода дополнительно включены слои возрастом 8–1.2 млн лет центральной и юго-восточной части Азии.
Отличительный диагноз
Предваряя описание отличительного диагноза необходимо отметить некоторые особенности и сложности.
Особенность диагностики объектов исследуемого периода филогенеза прямоходящих заключается в том, что мы не можем наблюдать вид – основатель рода, перешедший к прямохождению, а имеем дело с двумя, на которые он впоследствии распался и которые также находятся в состоянии последующих эволюционных преобразований. Настоящий диагноз должен не только путём ревизии старого материала и идентификации нового уточнить филогенез африканских прямоходящих периода 6.2–1.2 млн лет, но и показать путь восстановления «эволюционной непрерывности», вплоть до выявления протоморфного вида надсемейства гоминоиодов.
Эта особенность вынуждает автора сделать два допущения. Во-первых, предположить, что H. sapiens представляет собой прямоходящее существо, достигшее в общем русле эволюции органического мира определённого уровня биологической организации, и поэтому включить его терминальным биологическим видом в филетическую линию рода «прямоходящие», что позволит получить для сравнения признаки прямохождения в наиболее совершенном и завершённом виде. Во-вторых, предполагая, что миоценовый гоминоидный предок рода Bipedius и современные понгиды принадлежат в вертикальной палеонтологической систематике приматов к одному надсемейству гоминоидов, представляют экологические эквиваленты друг друга и сходны по способу локомоции, получить необходимые для сравнения признаки, диагностирующие «получетвероногую» исходную локомоцию.
Особой сложностью настоящего отличительного диагноза является необходимость индуктивного восстановления важнейших морфологических особенностей, в нашем случае это способность прямохождения, по весьма фрагментарным и, как правило, единичным костным остаткам некогда живущих зоологических объектов.
Только лишь два отдела костного скелета: тазовый пояс и голеностопный сустав, а также некоторые особенности морфологии черепа и лопатки позволяют облигатно свидетельствовать о принадлежности представителя надсемейства гоминоидов к роду Bipedius.
Череп. Естественная, сбалансированная посадка головы, позволяющая длинным осям орбит быть обращёнными вперёд, в отличии от подвешенной на затылочных мышцах головы гоминоидов, обеспечивается у двуногих прямоходящих морфологией области затылочного отверстия, расположенного в середине основания черепа:
– угол расположения затылочного отверстия относительно франкфуртской горизонтали для современных антропоидов варьирует от +19 до +45°, уменьшение этого угла до величин близких к нулю (у современного человека угол отрицательный и варьирует от -7 до -16 °) свидетельствует о сбалансированной посадке головы, свойственной прямохождению [Нестурх, 1970; Рогинский, 1977];
– в отдельных случаях дополнительным признаком прямохождения служит степень изгиба основания черепа, измеряемая величиной углов назион-базион-сфеноидале (точка на заднем крае sulcus chiacmaticus) так называемого треугольника основания черепа. Для антропоидов, у которых основание мало изогнуто, угол при сфеноидальной точке превышает 170°, у человека угол не превышает 150° [Рогинский, 1977];
– сосцевидный отросток (височной кости) у гоминоидов треугольный и плоский, у A. afarensis треугольный, но выпуклый, у человека овальный с глубокой вырезкой [Джохансон, Иди, 1984];
– морфология остистых отростков шейных позвонков, функционирующих у гоминоидов как аппарат уравновешивания относительно тяжёлого черепа, являясь местом прикрепления затылочной мускулатуры, позволяет, правда косвенно, судить о её мощности [Хрисанфова, 1978];
– также косвенно можно судить о мощности затылочной мускулатуры по рельефу чешуи затылочной кости, в частности по степени развития наивысшей, верхней и нижней выйной линии и степени выступания наружного затылочного выступа. Так, наружный затылочный гребень, характерный как для миоценовых, так и для современных гоминоидов, отсутствует даже у прямоходящих растительноядных, для которых, тем не менее, характерен саггитальный гребень для крепления жевательных мышц [Хрисанфова, 1978];
– шейный лордоз характерен для прямохождения, однако вертикальный указатель отдела не позволяет сделать никаких заключений о степени выраженности изгиба, поскольку лордоз обусловлен исключительно межпозвоночными дисками [Хрисанфова, 1978].
Тазовый пояс. Тазовый пояс играет особую роль в качестве индикатора бипедии. Его структура является чётким критерием разграничения двуногих прямоходящих надсемейства гоминоидов от остальных членов надсемейства.
Для прямоходящих рода Bipedius характерен следующий комплекс основных признаков тазовой кости: укорочение подвздошной кости, вследствии дорзальной ротации подвздошно-крестцовых сочленений по отношению к тазобедренным суставам, что обусловило и уменьшение расстояния между этими суставами; расширение пластинки крыла в дорзальном и вентральном направлении и укрепление её средней части; укрепление вертлужной впадины и изменение её положения; отчётливое оформление большой седалищной вырезки и обеих передних подвздошных остей; расширение ушковидной суставной поверхности, сочленяющейся с крестцовой костью:
– высотно-широтный указатель таза современных «антропоморфных» приблизительно составляет 90, для современного человека около 80 [Нестурх, 1970; Рогинский, 1977];
– степень отклонения назад подвздошной кости характеризуется углом, образуемым осями лобковой кости и крестцовой части подвздошной кости. Ось лобковой кости образуется линией, проведённой из центра суставной впадины к верхнему краю симфиза лобковых костей, ось крестцовой части подвздошной кости – линией, проведённой из того же центра к середине ушковидной поверхности. Этот угол у современных «антропоморфных» варьируется от 125 до 145°, у человека – 160°. В связи с указанным поворотом крестцовой части подвздошной кости у человека образуется глубокая большая седалищная вырезка [Рогинский, 1977];
– глубинно-широтный указатель таза (индекс большой седалищной вырезки) для антропоидов 13.5–22.2; для человека – 59.0–71.1; для A. africanus – 44.7 [Хрисанфова, 1978];
– относительное развитие вертлужной впадины и её положение является довольно устойчивым признаком прямохождения. Отношение диаметра вертлужной впадины к высоте таза составляет у человека 24.8–26.5; у A. africanus 24.6; у гоминоидов – 13.9–18.7 [Хрисанфова, 1978];
– следует упомянуть о наличии у прямоходящих поясничного лордоза, обеспечивающего статику или так называемую осанку прямоходящего, который представляет сложное образование, состоящее из нескольких компонентов, коррелирующих между собой. Остеометрические показатели, используемые при анализе ископаемых материалов, такие как вертикальный поясничный указатель и указатель клиновидности поясничного позвонка, не характеризуют полностью его степень, поскольку изгиб обусловлен в основном межпозвоночными дисками. Однако при вертикальном поясничном указателе ниже 98.0 всё же существует отчётливая выраженность поясничного лордоза [Хрисанфова, 1978];
– кроме остеометрических показателей большое значение для идентификации прямохождения имеет восстановление по косвенным признакам мускульного рельефа. Можно упомянуть лишь признак извилистости подвздошного гребня; перекладину на подвздошном гребне тазовой кости, отсутствующую у антропоидов, которая спускается от своей границы – передней верхней подвздошной ости – к тазобедренному суставу и является рычагом для мускулов, участвующих в балансировании туловища; мощный «вертикальный пилястр» на подвздошной кости; рельефы в местах прикрепления подвздошно-бедренной связки.
Голеностопный сустав. Прямохождение на стопе определяется, в первую очередь, комплексом «тибилизации», т. е. приведением и укреплением первого луча, принимающего большое участие в образовании опорной поверхности стопы, массивностью I плюсневой, пропорциями рычагов стопы, связанными с распределением статодинамической нагрузки, укреплением сводов [Хрисанфова, 1978]:
– для прямохождения характерно абсолютно и относительно сильное развитие первого пальца, который с одновременной утратой им способности противопоставляться остальным пальцам охвачен общей для всех пальцев стопы поперечной плюсневой связкой;
– суставная поверхность медиальной клиновидной кости, сочленяющейся с первой плюсневой, почти плоская и мало скошена в медиальном направлении, что стоит в связи с меньшей подвижностью и большим приведением большого пальца. Подвижность и склонность первой плюсневой кости к противопоставлению проверяется соответствием сочленяющихся поверхностей в первом клиновидно-плюсневом суставе и по участкам фиксации сухожилия (соответствующих бугристостей) длинной малоберцовой мышцы, сокращение которой вызывает противопоставление первого пальца стопы, а у человека, совместно с сокращением передней большеберцовой мышцы, способствует скручиванию стопы и углублению её поперечного свода [Данилова, 1989];
– в связи со значительной потерей функциональной роли II–V пальцев в опоре и локомоторной функции они у человека постепенно становятся на путь редукции, что в известной мере находит отражение и в форме соответствующих плюсневых костей: истончение дистальных отделов этих костей, уменьшение размеров и поперечная сдавленность их головок, уплощение выраженного суставного рельефа в клино-ладьевидном и предплюсне-плюсневых суставах [Данилова, 1989];
– таранная кость у человека сравнительно короткая, широкая и низкая с длинной и массивной шейкой. Характерной особенностью является меньшее, по сравнению с антропоморфными, отклонение шейки в сторону, что связано с приближением большого пальца к остальным; угол, образуемый осью шейки с осью тела кости, составляет у человека 20–25°, у антропоидов – выше 30° [Рогинский, 1977]; угол между осями таранной и пяточной кости у человека (от -8 до +12°) меньше чем у антропоидов (от +16 до +26°) [Нестурх, 1970];
– в связи со слабым отклонением шейки таранной кости, опора таранной кости на пяточной кости сравнительно узка, а угол, образуемый длинной осью задней суставной площадки с продольной осью кости, велик. У человека он равен в среднем 40–45°, у гоминоидов 5–15°. Суставная поверхность для сочленения с кубовидной костью повёрнута таким образом, что её ось приближается к вертикали, тогда как у гоминоидов она лежит почти в горизонтальной плоскости, что связано у человека с образованием поперечного свода;
– медиальное перемещение функциональной и морфологической оси стопы и связанная с этим перегруппировка межкостных мышц; расширение предплюсны – её ширина в процентах длины III луча равна у человека 99, у гориллы – 84, шимпанзе – 75 [Данилова, 1989];
– сводчатое строение составляет характерную особенность стопы человека. Под продольным сводом понимают пять продольных лучей, образующих дуги, которые идут от пяточного бугра к основаниям плюсневых костей; при этом медиальная часть стопы приподнята и при опоре не касается опорной поверхности. Продольный свод можно определить по форме и положению суставных площадок в пяточно-кубовидном суставе [Данилова, 1989];
– дугообразное взаиморасположение клиновидных и кубовидной костей стопы указывает на наличие выраженного поперечного свода, при этом длинная ось суставной поверхности головки таранной кости, сочленяющейся с ладьевидной костью, сильно отклонена от горизонтальной плоскости; угол, образованный этой осью с горизонталью (угол торзиона головки) составляет у человека 30–40°, у орангутанга – 10°, у гориллы – 20° (поперечный свод свойственен не только «прямоходящим») [Рогинский, 1977];
– пяточная кость отличается большой шириной и особенно сильным развитием бугра, на котором имеется хорошо выраженный боковой отросток (processus lateralis tuberus calcanei), составляющий специфическую особенность этой кости у человека;
– наличие контактных фасеток между основаниями I и II плюсневых костей позволяет судить о сравнительном уровне предплюсно-плюсневых суставов обоих большеберцовых лучей (отсутствие фасеток не говорит о противоположном).
Бедренная кость. Длинные кости нижних конечностей гоминоидов не показывают признаков, позволяющих, каждый в отдельности, облигатно индицировать прямохождение. Выделяемые морфологические признаки, как правило, имеют множественную природу и значение, причём существующая значительная трансгрессия значений остеологических показателей в строении длинных костей конечностей у прямоходящих и гоминоидов позволяет, в основном определить отсутствие «противопоказаний» в признании бипедии. Это имеет особое значение при фрагментарном состоянии материала и требует структурно-функционального анализа костей при привлечении не изолированных признаков, но их комплекса [Хрисанфова, 1978]:
– характерной особенностью бедренной кости у прямоходящих является сильное развитие шероховатой линии (linea aspera), которая образует в разной степени выраженный гребень "пилястр", определяющий усиление мускулатуры бедра, в том числе широких мышц (m. vastus lateralis, m. vastus medialis), которые берут начало от губ шероховатой линии. Однако пилястр может рассматриваться и как контрфорс, возникший в результате адаптивной реакции костной ткани на изгиб бедра (укрепление задней стенки). Указатель пилястрии характеризует форму сечения диафиза, взятого на уровне наибольшего развития шероховатой линии. У человекообразных обезьян шероховатая линия не развита, бедренная кость уплощена в переднезаднем направлении и указатель пилястрии соответственно низок: у гориллы – 75, орангутанга – 78, шимпанзе – 85, у современного человека он варьирует в пределах 100–122; у неандертальца – 100, у питекантропа – 109 [Рогинский, 1977];
– структуру диафиза бедренной кости отражает также весьма существенный для прямохождения признак скрученности кости – её торзион, определяемый взаимным расположением оси верхнего эпифиза (шейки) кости и касательной к мыщелкам нижнего эпифиза. Для человека типичен «положительный торзион», когда ось шейки проходит сзади латерально, вперёд медиально. Групповые вариации признака составляют 8–39.7. Для гоминоидов обычны отрицательные углы (ретроверсия) [Хрисанфова, 1978];

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
