

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
С. В. Вырский
Реконструкция
филогенеза «австралопитеков»
методом сравнительного анализа
признаков
Саратовский государственный университет им. Н. Г. Чернышевского
С. В. Вырский
Реконструкция
филогенеза «австралопитеков»
методом сравнительного анализа
признаков
Издательство Саратовского университета
2004
УДК 572.4
![]() ББК 28.71
ББК 28.71
В92
Вырский С. В.
В92 Реконструкция филогенеза «австралопитеков» методом сравнительного анализа признаков. – Саратов: Изд-во Сарат. ун-та, 2004. – 48 с.: ил.
ISBN 5–292–03163-1
Монография посвящена одной из наиболее актуальных тем современной палеоантропологии – филогенезу африканских прямоходящих приматов.
Последовательное применение таксономии и классификации позволяет автору не только выделить биологические виды, представленные ископаемыми остатками возрастом 8-1 млн лет, но и исследовать некоторые аспекты их видообразования.
Представленная систематика и классификация прямоходящих приматов, безусловно, будет интересна как специалистам в области палеоантропологии, так и широкому кругу биологов.
Ил. 2. Табл. 3. Библиогр.: 20 назв.
Рецензенты:
Доктор биологических наук, профессор Г. В. Шляхтин
Доктор геолого-минералогических наук, профессор В. Г. Очев
Работа издана в авторской редакции
УДК 572.4
ББК 28.71
 ISBN 5–292–03163-1 © Вырский С. В., 2004
ISBN 5–292–03163-1 © Вырский С. В., 2004
введение
 |
В настоящее время африканский континент находится в центре полевых и теоретических исследований, посвящённых австралопитекам. Сотни найденных и идентифицированных зоологических объектов, разнообразие видов, новые временные интервалы, из которых они получены, но одновременно и отсутствие концепций, связывающих воедино это многообразие, побудили автора вернуться к проблеме австралопитеков. Важной причиной написания настоящей работы явилось желание автора проверить соответствие модели их происхождения [Вырский, 1989] новому палеоантропологическому материалу. В данной работе автор исследовал филогенез австралопитеков, или, если следовать контексту монографии, филогенез африканских прямоходящих приматов «австралопитекового» периода.
Восстановление или реконструкция филогенеза проводилась применением стандартных методов классификации зоологических объектов, таксонированием набора ископаемых остатков, номинируемых австралопитеками, с последующим разделением на таксоны более низкого уровня с помощью метода сравнительного анализа признаков [Майр, 1971]. Единственным отличием представляется принцип, положенный автором в основу выбора признака таксонирования и который можно назвать «системным»: признак должен выделять таксон симпатрически, но сохраняться или прослеживаться на образцах аллохронно. Это позволило, во-первых, разделить имеющийся набор зоологических объектов на таксоны, в которых проявляются признаки вида и, во-вторых, систематизировать полученные виды, т. е. восстановить вертикальные, по осадочным слоям, или аллохронные филетические связи.
В результате неупорядоченный набор зоологических объектов был разделён на таксоны видового, с точки зрения биологической концепции вида, уровня, установлены их систематические связи, а сами таксоны были классифицированы с присвоением номенклатуры.
Автор выражает благодарность за поддержку д-ру геол.-минер. наук, проф. института геологии БФАН РФ г. П. Рождественскому; за критические замечания д-ру геол.-минер. наук, проф. геологического факультета СГУ В. Г. Очеву; за ценные замечания доц. кафедры антропологии СГМУ, канд. мед. наук Т. М. Загаровской, за помощь и поддержку руководству и сотрудникам биологического факультета и кафедры морфологии и экологии животных СГУ; проф. университета штата Аризона (США), директору I. H.O. д-ру Дональду С. Джохансону за предоставленные материалы; за оказанную техническую помощь при подготовке работы инженерам В. Шраму, А. Семёнову, О. Лонщаковой; переводчику английского языка Г. В. Алексееву, переводчику латинского языка Е. Ю. Кузнецовой.
 I. предварительное таксонирование
I. предварительное таксонирование
на основе Современных представлений
филогении австралопитеков
Предварительное таксонирование, необходимое для восстановления или реконструкции филогенеза австралопитеков, безусловно, следует начать с рассмотрения основных эволюционных этапов, признаваемых в настоящее время большинством исследователей.
Если обратить внимание на возраст слоёв, в которых в последнее время производился поиск костных остатков австралопитеков, то можно увидеть, что этот поиск происходит в двух направлениях, определяемых предполагаемыми узловыми моментами их филогенеза.
В основе первого направления исследований лежит предположение, что происхождение австралопитеков непосредственно связано с африканским континентом, в 1992–1994 гг. найден вид Australopithecus ramidus возрастом 4.4–4.2 млн лет [White et al., 1994]; в 2000 г. – вид Orrorin tugenensis возрастом 6.2–5.65 млн лет [Senut et al., 2001]; в 2001 г. вид Sahelanthropus tchadensis возрастом 7.0–6.0 млн лет [Brunet et al., 2002]. Фактически, исследования последовательно уходят в более глубокие временные слои с целью поиска костных остатков, связывающих австралопитеков с какой-либо из предшествующих форм надсемейства гоминоидов.
В основе второго направления исследований лежит факт симпатрического сосуществования на африканском континенте в интервале времени 2.5–1.2 млн лет австралопитеков и первых видов рода Homo (табл. 1). Учитывая, что все они относятся к прямоходящим, исследователи надеются найти их непосредственную взаимосвязь. Поиск в слоях этого интервала времени дал в 1996–1998 гг. вид Australopithecus garhi возрастом 3–2 млн лет [Asfaw et al., 1999] и в 1998–1999 гг. – вид Kenyanthropus platyops (3.5‑3.2 млн лет) [Leakey et al., 2001].
Эти два направления фактически свидетельствуют, что большинство современных полевых исследователей филогенеза австралопитеков, во всяком случае в виде рабочей схемы, рассматривают гоминоидов, австралопитеков и Homo как взаимопереходящие формы единой эволюционной линии.
Эти три формы – «гоминоиды – австралопитеки – Homo» взяты автором как три таксона, на которые предварительно разделён исследуемый набор зоологических объектов. Восстановление определительных признаков, на основании которых произведено это разделение, и последующее таксонирование, о чём говорилось во введении, будет предметом дальнейшей работы.
II. отделение прямоходящих австралопитеков
и HOMO от непрямоходящих – гоминоидов
Девять видов, номинируемых австралопитеками, показывают совокупный хронологический интервал распространения от 4.4 до 1.2 млн лет назад (см. табл. 1). Несомненно, наиболее общим, «генеральным» признаком, присущим всем этим видам и отличающим их от остальных членов надсемейства гоминоидов, которому предположительно они принадлежат, является признак «прямохождение». «Прямохождение» уверенно проверяется операционально на костных остатках в виде множества фенотипических признаков бедра, стопы, большого затылочного отверстия и т. д., является в этом смысле интегральным признаком, и автор считает возможным объединить все 9 видов австралопитеков в таксон, которому присвоить название одноимённое с таксоноопределяющим признаком – «прямоходящие».
Таблица 1
Перечень видов, включённых в таксон «прямоходящие»
№ п/п | Оригинальные виды | Время, млн лет | № п/п | Оригинальные виды | Время, млн лет |
1 | Orrorin tugenensis | 6.2–5.65 | 8 | Australopithecus garhi | 2.5 |
2 | Australopithecus ramidus | 4.4–4.2 | 9 | Australopithecus aethiopicus | 2.7–2.39 |
3 | Australopithecus anamensis | 4.17–3.9 | 10 | Australopithecus robustus | 2.0–1.5 |
4 | Australopithecus afarensis | 4.0–2.33 | 11 | Australopithecus boisei | 2.5–1.2 |
5 | Australopithecus bahrelghazali | 3.5–3.0 | 12 | Homo rudolfensis | 2.4–1.8 |
6 | Kenyanthropus platyops | 3.5–3.2 | 13 | Homo habilis | 2.3–1.5 |
7 | Australopithecus africanus | 3.3–2.3 | 14 | Homo ergaster | 1.8–1.2 |
Обратим внимание на то, что к этому интервалу времени относятся также 3 вида рода Homo – H. rudolfensis [Leakey , 1973], H. habilis [Leakey et al., 1964], H. ergaster [Groves and Mazak, 1975] и вид K. platyops [Leakey et al., 2001], которые также являются прямоходящими и по определению должны быть включены в этот таксон.
Ещё один вид – Orrorin tugenensis [Senut et al., 2001], хотя и относится к более раннему времени распространения (6.2–5.65 млн лет), также претендует на прямохождение и подлежит включению в таксон.
Нельзя не упомянуть хронологически самый ранний и потому представляющий несомненный интерес, вид Sahelanthropus tchadensis (7.0–6.0 млн лет) [Brunet et al., 2002], однако его способ передвижения ещё недостаточно исследован, что не позволяет в настоящее время включить его в образованный таксон.
Сформировав таксон «прямоходящие» и расположив таблично в хронологическом порядке объединённые в нём 14 видов (см. табл. 1), мы получаем первый важный вывод: прямохождение и соответственно аллохронный таксон «прямоходящие» сформировались не позднее 6.2–5.65 млн лет назад. Одним из доказательств этого служит найденная в этом временном интервале проксимальная часть левого бедра – образец BAR-1002'00 вида Orrorin tugenensis [Brunet et al., 2002].
III. расчленение таксона «прямоходящие»
с помощью признака «индекс энцефализации»
Признак «индекс энцефализации». Если границу между австралопитеками и предшествующими формами гоминоидов можно определить временем возникновения таксона «прямоходящие», то для нахождения границы филогенеза между собственно австралопитеками и Homo, внутри таксона «прямоходящие», необходим признак, который бы отражал уровень гоминизации. Таким признаком, уверенно проявляющимся на краниальных остатках образцов, является «индекс энцефализации».
Для расчёта индекса энцефализации (ИЭ) автор применил формулу «квадратного весового указателя мозга», или «отношение квадрата веса мозга к весу тела» [Рогинский, 1977]. Эта формула идентична «относительному весу мозга», но более удобна для отображения в прямоугольной системе координат:
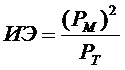 , (1)
, (1)
где: Pм – вес мозга, в г; Pт – вес тела, в г.
Поскольку костные образцы дают величину объёма черепной коробки, то для перехода к весу мозга принимаем следующую пропорцию: 100 см3 объёма мозгового вещества соответствуют 91 г веса.
Следует отметить, что объём головного мозга несколько меньше вместимости черепной коробки, однако в рамках данной работы это отклонение для ИЭ не имеет существенного значения.
Построение графика, отражающего изменения индекса энцефализации. Для подготовки данных, необходимых для расчёта ИЭ, автор составил перечень образцов, относящихся к видам, включенным в таксон «прямоходящие» [Heslip, 2001; Kreger, 2002] и расположил их таблично (см. прил.). Дополнив таблицу (см. прил.) отдельными характеристиками, необходимыми для расчёта ИЭ, и произведя расчёты величин ИЭ по формуле (1), автор обобщил полученные результаты, в виде областей значений интегрированных характеристик для различных видов, а также конкретные значения ИЭ для отдельных образцов (табл. 2).
Таблица 2
Значения индекса энцефализации для видов и отдельных образцов
таксона «прямоходящие»
№ п/п | Вид | Зоологический | Возраст, | Индекс энцефализации |
1 | Australopithecus afarensis | 4.0–2.75 | 4.8/3.4 | |
2 | Australopithecus afarensis | AL 288 «Lucy» | 3.2 | 3.8 |
3 | Australopithecus africanus | 3.0–2.5 | 5.1/3.5 | |
4 | Australopithecus boisei | 2.6–1.0 | 4.14/3.05 | |
5 | Australopithecus boisei | OH 5 «Zinj» | 1.8 | 3.88 |
6 | Homo habilis | 2.4–1.5 | 5.2/8.8 | |
7 | Homo habilis | OH 7 | 1.75 | 11.04(9.1/13.2) |
8 | Homo habilis | OH 13 «Cindy» | 1.66 | 5.52 |
9 | Australopithecus ramidus | 4.4 | 4.41 | |
10 | Australopithecus robustus | 2.0–1.0 | 3.88 | |
11 | Homo rudolfensis | 2.4–1.6 | 8.12 | |
12 | Homo ergaster | KNM WT 15000 | 1.6 | 12.74 |
13 | Orrorin tugenensis | 6.2–5.65 | 4.4 |
Безусловный интерес представляет также сравнение по ИЭ объектов таксона «прямоходящие» с предшествующими им формами азиатских или африканских гоминоидов, однако отсутствие хороших данных для последних не позволило рассчитать значения индекса. Тем не менее, полагая, что для современных человекообразных обезьян семейства Pongidae и для их экологических, локомоторных и систематических эквивалентов миоценового периода, входящих в надсемейство Hominoidea, из которых предположительно возникли австралопитеки, индекс будет примерно одинаков. Автор представил в табл. 3 области значений ИЭ для двух наиболее характерных видов современных человекообразных обезьян.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
