

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
– стабилен у человека кондилодиафизарный угол – угол инклинации, составленный анатомической осью кости и касательной к плоскости обоих мыщелков («коленному базису»), имеющий вариации 72–88° и, правда, некоторую трансгрессию с гоминоидами 79–94°;
– определённое значение для прямоходящих имеет удлинённая шейка бедренной кости и её коллодиафизарный угол 121–133°, определяющий положение шейки бедра по отношению к её диафизу, для современных антропоидов он составляет 109–138°;
– нижний эпифиз кости прямоходящих показывает преобладание трансверзального развития обоих мыщелков над саггитальным, а также относительно крупный наружный мыщелок;
– тенденция у человека к равенству площадей частей сочленовых поверхностей обоих мыщелков, контактирующих при полном разгибании колена с верхними суставными поверхностями большеберцовой кости. Процент отношения величины тибиальной поверхности наружного мыщелка к таковой внутреннего варьирует от 80.6–108.6 для человека и 57.7–70.9 для современных антропоидов [Хрисанфова, 1978];
– нижний конец бедренной кости имеет такой важный признак, как уплощённый контур мыщелков, указывающий на полное разгибание колена, в отличии от округлого контура мыщелков гоминоидов [Данилова, 1989];
– функциональная гипертрофия структуры длинных костей, связанная с усилением нагрузки на нижние конечности, проявляется в особенностях трабекулярной структуры проксимального эпифиза бедренной кости, в структуре диафизарных отделов бедренных костей, прежде всего кортико-медуллярных пропорций и соотношении компактного костного вещества и костномозговой полости. В компактном костном веществе эти процессы проявляются в его утолщении (при сужении костномозговой полости), крупноячеистости типа губчатого вещества, повышении остеонизации [Хрисанфова, 1978].
Берцовые кости. Верхний эпифиз большой берцовой кости несколько отогнут кзади, образуя с осью диафиза небольшой угол, называемый углом ретроверсии, измеряемым между осью диафиза и саггитальной прямой, проведённой через суставную поверхность медиального мыщелка верхнего конца большой берцовой кости. Этот угол у человека равен в среднем 12°, у «антропоморфных» – 25° [Рогинский, 1977]:
– ось верхнего эпифиза большой берцовой кости, проходящая через центры суставных поверхностей, образует с осью нижнего эпифиза (делящей его суставную площадку пополам) острый угол, называемый углом торзиона или скрученности. У современного человека это так называемый положительный угол торзиона в пределах от +6 до +33°; напротив, у «антропоморфных» угол торзиона отрицательный – от -7.5 до -24° [Рогинский, 1977; Хрисанфова, 1978].
Пояс верхних конечностей. В наибольшей степени эволюционные преобразования передних (грудных) конечностей прямоходящих, связанные с появлением и развитием разнообразных движений в трёхосном плечевом суставе, сказались на лопатке:
– уменьшение морфологической длины и увеличение морфологической ширины, связанное с перемещением лопаток из бокового положения на стенке грудной клетки у полуобезьян в дорзальное, с соответственным изменением положения суставной впадины;
– суставная впадина развёрнута у человека латерально и располагается, в общем, параллельно вертикальной оси лопатки, тогда как у других приматов сочленяющаяся поверхность повёрнута более или менее краниально. С поворотом суставной впадины связано и различное расположение лопаточной ости. У человека ость (spina scapulae) располагается почти трансверзально, у «человекообразных» – косо, а у «низших обезьян» – саггитально. Угол, образуемый лопаточной остью с вертикальной осью лопатки, равен в среднем: у человека – 88°; у гориллы – 59°; у орангутанга – 56°, у шимпанзе – 40°, у гиббона – 32°;
– аксиллогленоидальный угол, измеряемый между латеральным краем и плоскостью суставной впадины лопатки, составляет для антропоидов величину около 108°, для современного человека – 132° [Хрисанфова, 1978];
– аксиллярному (латеральному) краю лопатки антропоидов свойственна простая форма края.
Кисть. Прежде всего, необходимо указать на значительную примитивность кисти «прямоходящих» исследуемого периода. Сюда нужно отнести полуфлексированное положение пальцев, значительную расширенность диафизов проксимальных и средних фаланг, топографию и выраженность гребней для прикрепления поверхностного сгибателя пальцев, положение кости трапеции среди других костей запястья и морфологию головчатой кости. Кисть лишь слегка затронута процессом гоминизации, вероятно, более близкие к человеческим пропорции пясти и пальцев, уменьшение изогнутости фаланг, возможное относительное удлинение I луча и некоторое его смещение из плоскости остальных, сравнительно с соответствующими соотношениями на кисти гориллы [Хрисанфова, 1978].
Интегральным признаком, отличающим кисть прямоходящих от кисти гоминоидов, является сформированный кистевой замок, условно представляющий противопоставление I луча кисти и его волярное (ладонное) смещение из плоскости остальных лучей. При этом следует иметь в виду широкие компенсаторные возможности мускульно-связочного аппарата, о которых трудно судить на основании остеологических материалов.
Противопоставление I луча кисти:
– определённый уровень абсолютного и относительного развития I луча и его компонентов: индекс соотношения I - II лучей у современного человека составляет 65.3–66.1 у антропоидов 48.5–50.2 [Хрисанфова, 1978];
– положение и форма костных компонентов соответствующего запястно-пястного сустава. В частности, седловидная форма сустава отчётливо выражена на кости трапеции, где саггитальная выпуклость суставной поверхности комбинируется с её поперечной вогнутостью;
– увеличение конгруэнтности суставных поверхностей и их уплощение [Хрисанфова, 1978];
– расширение ногтевых фаланг. Широтно-длиннотный указатель дистальных фаланг I–III–IV пальцев для человека дают ряд 41.7–42.7–40.6; наиболее близкие по значениям указатели для гориллы дают 33.7–35.5–32.1;
– развитый рельеф в местах прикрепления длинного сгибателя большого пальца, дифференцировавшегося от общего глубокого сгибания пальцев.
Анатомической основой волярного смещения I луча из плоскости остальных лучей является радиальное отклонение трапеции и скошенность её дистальной суставной поверхности. Большая многоугольная и ладьевидная кости запястья также смещены в волярном направлении. У антропоидов также существует некоторая ладонная скошенность основания I пястной кости, однако отклонение трапеции носит проксимальный характер.
Дополнительные признаки прямохождения:
– интермембральный показатель (процентное отношение суммарной длины плечевой и лучевой кости к длине бедренной и большеберцовой) чётко дифференцирует «прямоходящих» от гоминоидов. Групповые вариации его у современных людей составляют 64.4–74.9, у антропоидов средние показатели варьируют в пределах: 106.3 (шимпанзегиббоны). Интермембральный показатель для A. africanus соответствует
75.4–85.7, что близко к верхнему пределу вариаций у современного человека [Хрисанфова, 1978];
– важным диагностическим критерием, дифференцирующим прямохождение, является плечебедренный указатель. У человека современного типа пределы его групповых вариаций составляют 68.8–72.9; у шимпанзе - 101.0; у орангутанга – 139.0;
– следует указать ещё лучеберцовый указатель, который варьирует у современных людей в пределах 62.6–70.9 против 112.0–169.2 у антропоидов [Хрисанфова, 1978].
2. Вид «Прямоходящие растительноядные»
Интегрированному признаку «растительноядная специализация», определяющему таксон «прямоходящие растительноядные», присвоить ранг видоспецифичного признака, самому таксону «прямоходящие растительноядные» присвоить ранг биологического вида, входящего в род Bipedius, вновь образованному виду присвоить название Bipedius phytophagus sp. nov.
Этимология
Слово «phytophagus» в переводе с латыни означает «растительноядный», что соответствует специализации по питанию и раскрывает значение видоопределяющего признака.
В состав вида «Bipedius phytophagus» включить оригинальный вид A. ramidus, образец KNM-KP 29281 оригинального вида A. anamensis, образец BOU-VP-17/1 оригинального вида A. garhi и оригинальные виды A. robustus, A. boisei.
Оригинальным видам, включённым в состав «B. phytophagus» и имеющим первоначальный ранг «вид», снизить ранг и присвоить статус «временной подвид» (temporal subspecies) с использованием триноминального названия (ст.5 и ст.45 Международного кодекса зоологической номенклатуры), т. е. с добавлением обозначения ключевого признака, данного при первоначальном обозначении образца. Таким образом, оригинальный вид A. ramidus переименовать в Bipedius phytophagus ramidus, образец KNM-KP 29281 оригинального вида A. anamensis переименовать в B. f. anamensis, оригинальный вид A. garhi переименовать в B. f. garhi, оригинальные виды A. robustus и A. boisei из области «IIIb» переименовать в B. f. robustus и B. f. boisei соответственно.
Что касается остальных образцов оригинальных видов A. anamensis и A. garhi, а также других оригинальных видов таксона «прямоходящие растительноядные» из области «IIIa», то само их включение в новый вид должно быть произведено после ревизии ископаемого материала, поскольку первоначальные названия давались на основе иных определительных признаков и в состав оригинальных видов иногда включались образцы, имеющие различные видоспецифичные, с точки зрения биологической концепции вида, признаки.
Описание вида.
Вид Bipedius phytophagus характеризуется морфологией, определяемой необратимой адаптацией к растительноядности. На костных остатках это проявляется в первую очередь в виде жевательной и дробящей специализации зубочелюстного аппарата.
Ареал вида.
Африканский континент.
Горизонты залегания.
Возраст слоёв залегания определён во временном интервале 4.4 –1.2 млн лет.
Типовой образец.
Типовым образцом вида «Bipedius phytophagus» назначается образец «OH-5» временного подвида B. f. boisei (оригинальный вид A. boisei) возрастом 1.8 млн лет, найденный Мэри Лики в 1959 г. в Олдувае Танзании и названный «Zinjanthropus». Данный образец представляет заключительную фазу развития растительноядной специализации и в этом смысле служит эталонным носителем набора видоспецифичных признаков, отличающих вид от материнского вида.
3. Вид «Прямоходящие всеядные»
Интегрированному признаку «всеядная специализация», определяющему таксон «прямоходящие всеядные», присвоить ранг видоспецифичного признака, самому таксону «IIIa», входящему в таксон «прямоходящие всеядные», присвоить ранг биологического вида, входящего в род «Bipedius», вновь образованному виду присвоить название Bipedius omnivorus sp. nov.
Этимология.
Слово «omnivorus» в переводе с латыни означает всеядный, что соответствует специализации по питанию и раскрывает значение видоопределяющего признака.
В состав вида «Bipedius omnivorus» включить членов таксона «IIIa» – два образца Bar 1001'00 и BAR 1425'00 оригинального вида Orrorin tugenensis и образец AL 200-1a оригинального вида A. afarensis.
Оригинальным видам, включённым в состав «B. omnivorus» и имеющим первоначальный ранг «вид», снизить ранг и присвоить статус «временной подвид» (temporal subspecies) с использованием триноминального названия (ст.5 и ст.45 Международного кодекса зоологической номенклатуры), т. е. с добавлением обозначения ключевого признака, данного при первоначальном обозначении образца.
Таким образом, образцам Bar 1001'00 и BAR 1425'00 оригинального вида Orrorin tugenensis присвоить название Bipedius omnivorus tugenensis, образцу AL 200-1a вида A. afarensis присвоить название B. o. afarensis.
Описание вида.
Вид Bipedius omnivorus характеризуется морфологией, определяемой адаптацией к всеядности, и предполагает наличие мясной пищи в рационе.
На костных остатках всеядность проявляется виде альвеолярного прогнатизма, обеспечивающего «скусывающую» функцию – псалидонтность. Соответственно этому обнаруживается наличие лингвального износа верхних резцов и клыков и лабиального износа нижних. Увеличенный клык, выступающий из зубного ряда, наличие диастем, увеличенный надглазничный валик, свидетельствующий о значительной доли мясной пищи в рационе. Саггитальный гребень у всеядных отсутствует.
Ареал вида.
Африканский континент.
Горизонты залегания.
Слои залегания определены во временном интервале 6.2–2.5 млн лет.
Типовой образец.
Типовым образцом вида Bipedius omnivorus назначается гиподигма B. o. tugenensis. (оригинальный вид Orrorin tugenensis) возрастом 6.2 – 5.65 млн лет, найденный Senut и др. в Tugen Hills Кении в 2000 г. Признак всеядной специализации, носителем которой является образец, в сочетании с глубоким возрастом идентифицированного прямохождения, предполагает хорошую аналогию с ещё более ранними образцами этого вида, совершившими инвазию в эту экологическую нишу.
Отличительный диагноз.
Некоторое упрощение дифференциации состоит в том, что род «Bipedius», в исследуемом геохронологическом интервале, представлен только двумя биологическими видами – B. phytophagus и B. omnivorus, что позволяет использовать для идентификации образцов метод альтернативного исключения.
Всеядности характерен альвеолярный прогнатизм, обеспечивающий «скусывающую» функцию – псалидонтизм, что в первую очередь проявляется в различной форме альвеолярной дуги верхней и нижней челюсти, а также форме и износе зубов:
– параболическая форма альвеолярной дуги верхней челюсти, гиперболическая – нижней;
– увеличенный клык, выступающий из зубного ряда, наличие диастем;
– наличие лингвального износа верхних резцов и клыков и лабиального износа нижних.
У растительноядных псалидонтность отсутствует, для них характерны перетирающие жевательные движения, что проявляется в одинаковой форме альвеолярной дуги верхней и нижней челюсти, а также в износе зубов переднего отдела в окклюзивной плоскости:
– мелкие резцы, небольшой клык, функционально близкий к премолярам, моляризованные нижние премоляры, очень крупные моляры;
– наличие саггитального гребня на черепе, служащего для крепления мощных боковых жевательных мышц (отсутствие саггитального гребня не говорит об обратном);
– у мегагнатных форм завершающей стадии эволюции наблюдается молярный мегадонтизм (утолщение эмали) и особое строение мыщелков, позволяющее производить жевательные движения нижней челюстью в горизонтальной плоскости.
Следует упомянуть ещё о следующих возможных отличиях:
– шагающей походкой, при которой развит механизм вращательных движений таза, обладают только всеядные;
– отличие внутренней структуры костей, которая непосредственно связана с особенностями питания;
– различное соотношение изотопов углерода 13C и 12C в эмали зубов, являющееся устойчивым признаком участия зоологического объекта в различных эколого-энергетических цепочках [Sponheimer and Lee-Thorp, 1999]
4. Род «Homo»
Признаку «величина индекса энцефализации выше 5.15», приобретённому видом «Bipedius omnivorus» в процессе эволюции, присвоить ранг родоопределяющего признака. Вновь назначенному роду присвоить номенклатурное название «Homo».
Вновь образованный род включить в зоологическую номенклатуру отряда приматов следующим образом.
Палеонтологическая систематика:
· Отряд Primates Linnaeus, 1758
- Подотряд Anthropoidea Mivart, 1864 Надсемейство Hominoidea Gray, 1825 Род Bipedius, 2004 Вид Bipedius omnivorus, 2004 Homo, gen. nov., 2004, ex sp. Bipedius omnivorus
Этимология.
Слово «Homo» в переводе с латинского языка означает «человек». В привычном биологическом смысле присваивается таксонам при наличии у них какого-либо качественного признака, облигатно отличающего в сторону гоминизации от таксонов надсемейства гоминоидов. В этом контексте слово Homo представляет хорошо известное название, которому автор присвоил вначале конкретное биологическое, а затем соответствующее номенклатурное значение.
В состав рода Homo включить членов таксона/вида с рабочим названием «IIIc».
Описание рода Homo.
Род Homo характеризуется морфологией, определяемой двумя систематическими признаками, присущими материнскому виду, которые отражены в его названии Bipedius omnivorus, и эволюционно приобретённый признак «величина индекса энцефализации выше 5.15», уровень которого соответствует появлению изготовленных орудий труда.
Ареал рода.
Африканский континент.
Горизонты залегания.
Слои залегания определены для исследуемого периода в интервале 2.5–1.2 млн лет назад.
5. Вид «Homo manuscirens»
Признаку «величина индекса энцефализации выше 5.15», имеющему классификационный родовой признак, присвоить ранг видоспецифичного в контексте биологической концепции вида. Таксону «IIIc», имеющему этот признак и входящему в таксон «прямоходящие всеядные», присвоить ранг биологического вида, входящего в род Homo. Вновь образованному виду присвоить название Homo manuscirens sp. nov.
Этимология.
Слово «manuscirens» в переводе с латыни означает «умеющий руками», что отражает способность изготавливать орудия труда.
В состав вида включить оригинальные виды H. rudolfensis, H. habilis и H. ergaster, входящие в состав таксона «IIIc».
Оригинальным видам, включённым в состав «Bipedius omnivorus» и имеющим первоначальный ранг «вид», снизить ранг и присвоить статус «временной подвид» (temporal subspecies) с использованием триноминального названия (ст.5 и ст.45 Международного кодекса зоологической номенклатуры), т. е. с добавлением обозначения ключевого признака, данного при первоначальном обозначении образца.
Таким образом, оригинальному виду Homo rudolfensis присвоить название Homo manuscirens rudolfensis, оригинальному виду Homo habilis присвоить название Homo manuscirens habilis и оригинальному виду Homo ergaster присвоить название Homo manuscirens ergaster.
Описание вида.
Вид Homo manuscirens характеризуется той же морфологией, что и род Homo, основателем которого он является и который единственно входит в этот род в исследуемом интервале.
Ареал вида.
Африканский континент
Горизонты залегания.
Слои залегания для исследуемого периода определены в интервале
2.5–1.2 млн лет.
Типовым образцом нового вида Homo manuscirens назначается образец OH-7 (оригинальный вид H. habilis), один из немногих позволяющих говорить о возможной взаимосвязи черепа и посткраниальных остатков и соответственно о возможности расчёта ИЭ.
Отличительный диагноз.
Тот же, что и у рода Homo.
6. Обобщение представленной классификации
Представленная в настоящей работе вертикальная палеонтологическая классификация для исследуемого геохронологического интервала выглядит следующим образом:
Bipedius gen.
B. phytophagus 4.4–1.2 млн лет
B. omnivorus 6.2–2.5 млн лет.
Homo gen. ex sp. B. omnivorus
Homo manuscirens 2.5–1.2 млн лет.
Результаты реконструкции филогенеза африканских прямоходящих приматов периода 6.2–1.2 млн лет, полученные в настоящей работе и объединённые с результатами реконструкции филогенеза прямоходящих периода 8–5 млн лет [Вырский, 1989], можно представить в виде кладограммы (рис.2), где B. a-is представляет рабочее, неноменклатурное название азиатского вида, основателя рода Bipedius, первым перешедшего к прямохождению.
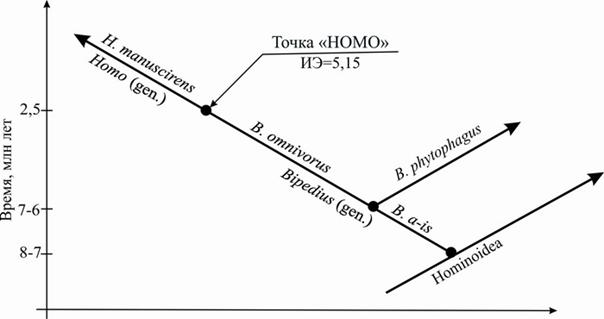
Рис. 2. Вертикальная палеонтологическая систематика прямоходящих приматов
периода 8–1.2 млн лет
ЗАКЛЮЧЕНИЕ
1. В настоящем исследовании произведена реконструкция филогенеза африканских прямоходящих приматов из временного интервала 6.2‑1.2 млн лет назад, номинированных ранее в основном как «австралопитеки». Выявлены и систематизированы, с точки зрения биологической концепции, и классифицированы с присвоением номенклатуры их виды.
2. Исследование выявило временные интервалы и признаки, представляющие наиболее важные, с точки зрения биологической концепции вида, моменты филогенеза, отражающие «незавершённое» видообразование, характеризуемое переходом популяций в иные эколого-энергетические ниши биоценоза, в частности переход к прямохождению, разветвление на два вида, специализированных по различным видам питания, переход к трудовой деятельности.
3. Исследование не опровергло модели филогенеза «австралопитеков», предложенной в работе [Вырский, 1989], однако значительно уменьшило предполагаемое время формирования морфотипа вида B. a-is, первым перешедшего к прямохождению, и/или расчётную скорость адаптивной радиации, т. е. времени необходимого для распространения от места возникновения на Лёссовом плато Центральной Азии до Африки. Время проникновения на африканский континент вида B. Omnivorus, указанное в монографии как 6-4 млн лет, должно быть увеличено по крайней мере до 6.2 млн лет. Время появления в Африке вида B. phytophagus, указанное как 4-3 млн лет, должно быть увеличено до величины не менее 4.4 млн лет назад.
4. Автор полагает, что последующая ревизия уже описанных образцов и видов, а также поиск новых костных остатков, в соответствии с представленной и уточнённой моделью филогенеза, позволит в ближайшее время завершить исследования филогенеза прямоходящих приматов периода 8-1.2 млн лет, окончательно систематизировать и классифицировать их виды, а также инструментальными методами определить фактическую модель видообразования для приматов, проверив также и концепции, стоящие в основании тех или иных моделей.
Библиографический список
Вырский С. В. К вопросу о происхождении австралопитеков. Саратов: Изд-во Сарат. ун-та, 1989. 20 с.
Данилова Е. И. Элементы гоминизации и дегоминизации в конечностях ископаемых и современных высших приматов // Биологическая эволюция и человек / Под ред. Я. Я. Рогинского. М.: МГУ, 1989. С. 104–124.
Джохансон Д. С., Иди М. А. Люси: истоки рода человеческого. М.: Мир, 19 с.
Майр Э. Принципы зоологической систематики. М.: МИР, 19 с.
Нестурх М. Ф. Происхождение человека. М.: Наука, 19 с.
Рогинский Я. Я. Проблемы антропогенеза. М.: Высшая школа, 19 с.
Хрисанфова Е. Н. Эволюционная морфология скелета человека. М.: МГУ, 19 с.
Asfaw B., White T. D., Lovejoy O. et al. Australopithecus garhi: A new species of early hominid from Ethiopia // Science. 1999. V. 284, № 5414. p. 629–635.
Brunet M., Guy F., Pilbeam D. et al. A new hominid from the Upper Miocene of Chad, Central Africa // Nature. 2002. V. 418. p. 145–151.
Coffing K., Feibel C., Leakey M., Walker A. Four-million-year-old hominids from east Lake Turkana, Kenya // American Journal of Physical Anthropology. 1994. v. 93. p. 55–65.
Groves C. P., Mazak V. An approach to the taxonomy of the Hominidae: Gracile Villafranchian hominids of Africa // Casopis pro Mineralogii a Geologii. 1975. v.20. p. 225–247.
Heslip S. J. Hominid fossils time and space chart. http://web. msu. edu/~heslipst/, Updated Spring Semester 2001.
Kreger D. A Look at Modern Human Origins. Hominid Index. http://www. /, Updated 28.09.2002.
Leakey L. S. B., Tobias P. V., Napier J. R. A new species of genus Homo from Olduvai Gorge // Nature. 1964. v. 202. p. 7–9.
Leakey M. G., Spoor F., Brown F. et al. New hominin genus from eastern Africa shows diverse middle Pliocene lineages // Nature. 2001. vol. 410. p.433–440.
Leakey R. E. Evidence for an advanced Plio-Pleistocene hominid from East Rudolf, Kenya // Nature. 1973. v. 242. p. 447–450.
Senut B., Pickford M., Gommery D. et al. First hominid from the Miocene (Lukeino Formation, Kenya) / C. R. Acad. Sci. Paris, Sciences de la Terre et des planets // Earth and Planetary Sciences. 2001. v. 332. p. 137–144.
Sponheimer M., Lee-Thorp J. Isotopic Evidence for the Diet of an Early Hominid, Australopithecus africanus // Science. 1999. Jan 15. Р. 368-370.
White T., Suwa G., Asfaw B. Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia // Nature. 1994. v. 371. p. 306–312.
Оглавление
Введение...................................................................................................... 3
I. Предварительное таксонирование на основе современных
представлений филогении австралопитеков................................. 5
II. Отделение прямоходящих австралопитеков и HOMO
от непрямоходящих – гоминоидов................................................... 6
III. Расчленение таксона «прямоходящие» с помощью признака
«индекс энцефализации» .................................................................. 7
IV. Расчленение таксона «прямоходящие»
по специализации в диете.............................................................. 11
V. Систематика прямоходящих периода 6.2–1.2 млн лет назад............ 13
VI. Классификация африканских прямоходящих приматов
периода 6.2–1.2 млн лет.................................................................. 16
Заключение................................................................................................. 32
Библиографический список ....................................................................... 33
Приложение........................................................................................ 34

Научное издание
Реконструкция филогенеза «австралопитеков»
методом сравнительного анализа признаков
Ответственный за выпуск Е. В. Завьялов
Технический редактор
Корректор
Подписано в печать.01.2004. Формат 60 х 84 1/16. Бумага офсетная. Гарнитура Таймс.
Печать офсетная. Усл. печ. л. 2,79(3). Уч.-изд. л. 2,8. Тираж 100. Заказ.
Издательство Саратовского университета. Саратов, Астраханская, 83.
Типография Издательства Саратовского университета. Саратов, Астраханская, 83.



|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
