
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На численность беспозвоночных в горных биоценозах достоверное влияние оказывают следующие факторы (в порядке убывания): «горно-растительный пояс» (F=8.1, p=0.0005) при min значениях средней численности в арктической пустыне и max в альпийском поясе → «гора» (F=5.2, p=0.02), min на г. Ловчорр, max на г. Поачвумчорр → высота над уровнем моря (F=3.7, p=0.0003), min на высоте 1093 м, max на высотах 590-600 м. На уровень биомассы влияет фактор «гора» (F=10.3, p=0.0001), min на г. Ловчорр, max на г. Поачвумчорр → «экспозиция склона» (F=4.2, p=0.01), min на северных склонах, max на южных.
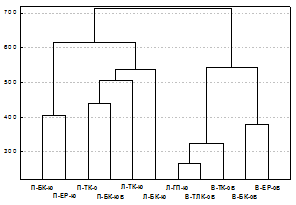
Ward`s method, Euclidean distances, 32 variants | Рис. 2. Дендрограмма фаунистического сходства горных и зональных биоценозов по разнообразию таксонов и численности почвенной фауны. Зональная тундра: ДЗ1-3, ДЗ-Б – окрестности пос. Д. Зеленцы, Т1-3, Т-ГТ – окрестности пос. Териберка; обозначение остальных биоценозов – как в табл. 1 и на рис. 1. |
ГЛАВА 7. ВИДОВОЕ РАЗНООБРАЗИЕ БЕСПОЗВОНОЧНЫХ В ПОЧВАХ ХИБИНСКИХ ГОР
Оценено разнообразие ключевых сапрофагов – дождевых червей и жуков трех семейств (стафилиниды, жужелицы, щелкуны), наиболее массовых в экосистемах гор.
7.1. Жесткокрылые (Coleoptera: Staphylinidae, Carabidae, Elateridae)
7.1.1. Общая характеристика разнообразия доминирующих семейств
Методами почвенных проб и ловушек выявлено 79 видов: 47 видов стафилинид из 27 родов, 7 п/сем.; 22 вида жужелиц из 13 родов, 4 п/сем.; 10 видов щелкунов из 10 родов, 4 п/сем. Впервые для Хибин указываются 42 вида стафилинид, 11 видов жужелиц и 6 видов щелкунов, впервые для Мурманской области – 19, 4 и 1 вид из этих семейств.
Наименее разнообразна фауна жуков г. Вудъяврчорр (30 видов: стафилинид 18, жужелиц 8, щелкунов 4 вида) из-за расположения большинства биоценозов на северо-восточном склоне. На г. Поачвумчорр и Ловчорр выявлено 54 и 46 видов жуков. Стафилиниды были наиболее разнообразны в лесных биоценозах г. Поачвумчорр: 21-24 вида из 13-15 родов (всего 33 вида, 18 родов). Жужелицы – на северных склонах: в субальпийском поясе г. Ловчорр (14 видов из 10 родов) и в тундровых биоценозах г. Поачвумчорр (10 видов из 7 родов), как и щелкуны (по 6 видов из 6 родов в каждом из этих биоценозов). Во всех горных экосистемах видовой спектр стафилинид был шире по сравнению с жуками других семейств. Общими для 3 гор были лишь 13 из 79 видов жуков (или 17%): по 5 видов стафилинид и жужелиц и 3 вида щелкунов.
Фаунистическое своеобразие жесткокрылых и, следовательно, различие экологических условий на разных горах подтверждено низкими значениями коэфф. Съеренсена (<22%) и результатами кластеризации данных (рис. 3). В один кластер (справа) выделились биоценозы северо-восточного склона г. Вудъяврчорр, в другой кластер - биоценозы гор Ловчорр и Поачвумчорр, расположенные в основном на склонах, ориентированных к югу. В пределах этих кластеров обособились горно-лесные биоценозы соседних поясов каждой горы и биоценозы субальпийского и альпийского поясов, что свидетельствует о взаимном проникновении таежных и тундровых видов жуков в экосистемы этих поясов.
| Рис. 3. Убывание сходства видового состава жесткокрылых (суммарно по трем семействам) в ряду: соседние горно-растительные пояса на склоне одной горы (коэф. Съеренсена 38-49%) → одноименные горно-растительные пояса на разных горах (7-36%) → одноименные горно-растительные пояса на разных склонах одной горы (13-15%). |
Для 14 биоценозов Хибин ни одного общего вида жуков из трех семейств не выявлено. Из 79 видов 46 (или 58%) встречались локально – лишь в 1-2 горных биоценозах, в т. ч. 29 видов (или 60%) стафилинид, 12 видов (55%) жужелиц, 5 видов (50%) щелкунов. Около половины всех жуков (31 вид или 39%), в т. ч. 16 видов стафилинид, 10 видов жужелиц и 5 видов щелкунов были приурочены к определенному растительному поясу. Наибольшее число видов каждого семейства (стафилинид 10, жужелиц 5, щелкунов 4) как лесной, так и тундровой преференции, населяли переходный субальпийский пояс, что можно рассматривать как проявление экотонного эффекта.
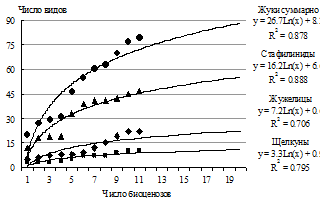
С учетом сильной расчлененности горного ландшафта и гетерогенности местообитаний, определяющих локальную встречаемость видов, можно прогнозировать обнаружение новых видов жуков. Кривые зависимости насыщения видовых списков от числа исследованных биоценозов [Коросов, 2007] показывают, что обследование 17 биоценозов (т. е. 5 гор) позволит учесть 99% видового разнообразия 3 семейств, что составляет около 90 видов (рис. 4).
| Число | ||
гор | биоценозов | видов | % |
3 | 11 | 79 | 94 |
4 | 14 | 84 | 98 |
5 | 17 | 88 | 99 |
Рис. 4. Увеличение числа видов жуков в горах с увеличением числа обследованных биоценозов. |
7.1.2. Семейство Стафилиниды (Staphylinidae)
В почвах трех гор выявлено 47 видов стафилинид из 27 родов, 7 п/сем. Наиболее разнообразны п/сем. Aleocharinae (19 видов) и Tachyporinae (10), адаптированные к обитанию в почвах северной тайги, тундры и горно-альпийских экосистемах [Колесникова, Ужакина, 2005]. По числу видов преобладали лишь 4 рода: Atheta (7 видов), Quedius (4), Oxypoda и Tachinus (по 3). Большинство родов (23 рода или 85%) включали по 1-2 вида, что отражает миграционный характер формирования горной фауны. По численности доминировали виды п/сем. Xantholininae и Aleocharinae: Xantholinus linearis, Othius lapidicola, Atheta brunneipennis, Drusilla canaliculata, Devia prospera. Доминирование отдельных видов в биоценозах горно-таежного и альпийского поясов не превысила 30% при разнообразии 12-20 видов. В экосистемах субальпийского пояса на фоне высокого числа видов (до 24) обилие эвритопного D. canaliculata достигало 40%.
Установлено влияние поясности на видовой состав, зоогеографическую и экологическую структуру комплексов стафилинид. Лесные по биотопическому преферендуму Deliphrum tectum, Atheta myrmecobia, Liogluta granigera, Liogluta micans, Oxypoda skalitzkyi и O. soror встречены лишь в таежном поясе гор. Еще 11 лесных, луговых и эвритопных видов обитали в горно-лесных поясах, не проникая в тундры. Только в субальпийских березняках выявлены 10 видов стафилинид лесной и тундровой преференции; только в альпийском поясе – лесные и тундровые Haploglossa villosula, Mniusa incrassata, Acidota quadrata, Anthophagus alpinus. В каменистом субстрате суровой арктической пустыни г. Ловчорр на высоте 1100 м обнаружены 4 вида, 3 из которых Atheta fungi, A. brunneipennis и Euaesthetus sp. являются новыми для Хибин, а вид Mycetoporus longulus, известный для высокогорных экосистем мира – новым для Мурманской области.
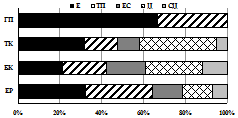
В целом сообщества этих жуков наиболее разнообразны в лесных поясах Хибин (27-34 вида против 4-18 в тундровых) и наиболее сбалансированы по зоогеографической и экологической структуре в субальпийском поясе (рис. 5).
Для Хибин было известно 15 видов стафилинид [Россолимо, 1994], из них нами не обнаружены 9 видов. С учетом всех данных в горных почвах обитает не менее 56 видов из 28 родов, 7 п/сем.. По числу видов преобладают рода Atheta (7 видов), Quedius, Oxypoda и Tachinus (по 3-4). Фауна стафилинид Мурманской обл. насчитывает 187 видов из 64 родов и 12 подсемейств при наибольшем разнообразии родов Stenus (23 вида), Atheta (15), Quedius, Philonthus (12-13) и Lathrobium (9).
I |
II |
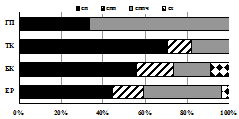
III | Рис. 5. Зоогеографическая (I), трофическая (II) и морфо-экологическая (III) структура комплексов стафилинид в высотных поясах Хибин. I: Ц – циркумареал, СЦ – субциркумареал, ТП – транспалеарктический, ЕС – европейско-сибирский, Е – европейский. II: с – сапрофаг, з – зоофаг, зм – зоомицетофаг, фд – фитодетритикол. III: сх – стратохортобионт, сп – стратобионт подстилочный, спп - стратобионт поверхностно-подстилочный, сппч – стратобионт подстилочно-почвенный. |
7.1.3. Семейство Жужелицы (Carabidae)
Выявлено 22 вида жужелиц из 13 родов 4 п/сем. Новыми для Хибин и региона в целом оказались 4 вида: Общими для трех гор были лишь 5 видов: Amara brunnea, Calathus melanocephalus, C. micropterus, Pterostichus brevicornis, Patrobus assimilis. Они встречались как в лесных, так и тундровых поясах Хибин. Максимальное число видов (10-14) характерно для альпийского пояса г. Поачвумчорр и субальпийского пояса г. Ловчорр. Наименьшее разнообразие (1-3 вида) отмечено в верхнем поясе арктической тундры г. Вудъяврчорр, полярной пустыне г. Ловчорр и лесных поясах южного склона г. Поачвумчорр. Наиболее высотное распространение в Хибинах (1093 м над ур. м.) установлено для жужелиц A. brunnea и P. brevicornis. Оба вида имеют обширные голарктические ареалы, а в Хибинах обитают в субальпийских и альпийских поясах всех гор.
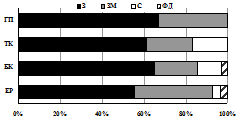
Изменению числа видов жужелиц в высотно-поясном градиенте (5→16→15→2) соответствует смена зоогеографической и экологической структуры их сообществ (рис. 6). Большинство видов, обитающих в лесных и тундровых поясах Хибин, являются зоофагами по типу питания и принадлежат к жизненной форме подстилочных или поверхностно-подстилочных стратобионтов. С высотой наблюдается смена преобладающей жизненной формы в ряду: подстилочные стратобионты → поверхностно-подстилочные стратобионты → геохортобионты. Хищные жужелицы вытесняются видами-фитофагами. Из 7 видов, встреченных в наиболее высотных поясах, 5 видов жужелиц в стадии имаго являются миксофитофагами, способными обитать в ярусе напочвенной растительности. Наиболее разнообразные сообщества жужелиц при сбалансированном соотношении зоогеографических и экологических групп характерны для субальпийских и альпийских поясов Хибин.
I |
II |
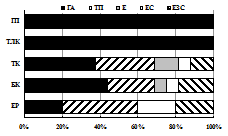
III | Рис. 6. Зоогеографическая (I), трофическая (II) и морфо-экологическая (III) структура комплексов жужелиц в высотных поясах Хибин. I: ГА – голарктический, ТП – транспалеарктический, Е – европейский, ЕС – европейско-сибирский, ЕЗС – Европейско-западносибирский; II: пс – подстилочный стратобионт, ппс – поверхностно-подстилочный стратобионт, эх (к) – эпигеобионт ходящий (крупный), гр – геобионт роющий, с – стратобионт, гх-г – геохортобионт гарпалоидный. III: з – зоофаг, м – миксофитофаг. |
В исследованных биоценозах трех гор нами не были обнаружены 6 видов жужелиц из 8 родов, указанные для Хибин в предыдущих работах [Россолимо, 1989, 1994; Эколого-экономическое обоснование…, 1999]; с их учетом карабидофауна Хибин насчитывает не менее 30 видов из 17 родов, а Мурманской обл. – 69 видов жужелиц из 26 родов относящихся к 8 п/сем. при наибольшем разнообразии родов Amara (17 видов), Bembidion и Agonum (по 7). В Хибинах по числу видов преобладают мелкоразмерные подстилочные хищники рода Notiophilus и травоядные жужелицы мелких и средних размеров из рода Amara (по 4 вида).
В исследованных биоценозах трех гор нами не были обнаружены 6 видов жужелиц из 8 родов, указанные для Хибин в предыдущих работах [Россолимо, 1989, 1994; Эколого-экономическое обоснование …, 1999]: Amara (=Paracelia) quenseli (Schönherr, 1806), Bembidion bipunctatum (Linnaeus, 1761), B. yukonum (Fall, 1926), Bradycellus caucasicus (Chaudoir, 1846), Cymindis vaporariorum (L., 1758), Elaphrus riparius (L., 1758), Loricera pilicornis (Fabricius, 1755). С учетом этих видов карабидофауна Хибин насчитывает не менее 30 видов из 17 родов, а Мурманской области в целом – 69 видов жужелиц из 26 родов относящихся к 8 подсемействам при наибольшем видовом разнообразии родов Amara (17 видов), Bembidion и Agonum (по 7). В Хибинах по числу видов наиболее разнообразны мелкоразмерные подстилочные хищники рода Notiophilus и травоядные жужелицы мелких и средних размеров из рода Amara (по 4 вида).
7.1.4. Семейство Щелкуны (Elateridae)
В биоценозах Хибин идентифицировано 10 видов щелкунов из 10 родов, 7 из них относятся к всесветно распространенному п/сем. Athoinae, разнообразному в горных экосистемах. Еще 3 вида родов Oedostethus и Limonius, предположительно виды O. nubilus, O. curticornis и L. parallelus, отловленные почвенными ловушками по 1 экз. в субальпийских поясах гор Ловчорр и Поачвумчорр, требуют уточнения. Щелкуны, ранее известные для Мурманской обл., но впервые выявленные в Хибинах, также отловлены в субальпийских поясах – березняке травянистом на южном склоне г. Поачвумчорр (лесные Mosotalesus impressus и Dalopius marginatus) и редкостойном березняке на северном склоне г. Ловчорр (борео-монтанный Orithales serraticornis и аркто-борео-монтанный Hypnoidus rivularius). В этом же березняке обнаружены мелкие гигрофильные: щелкун северный Ascoliocerus (Berninelsonius) hyperboreus, широко распространенный в зональных и горных тундрах Евразии, но новый для региональной фауны, и щелкун прибрежный Negastrius pulchellus, известный в Хибинах из работ [1936], но не найденный в других районах Мурманской обл.
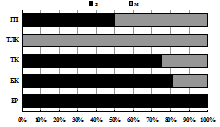
Аркто-альпийские и аркто-борео-монтанные N. pulchellus, A. hyperboreus и H. rivularius, способные выдержать суровые условия перигляциальной зоны, считаются пионерными видами щелкунов на северных территориях после отступления ледников [Медведев, 2005]. Бореальные и температные виды, населяющие горно-лесные пояса Хибин и составляющие основу региональной фауны элатерид, распространялись на север по мере потепления климата и появления лесов по интразональным биотопам из Европы и Сибири. Таковы борео-монтанные трансевразиатские Liotrichus affinis, Orithales serraticornis, Mosotalesus impressus, температный трансевразиатский Sericus brunneus, виды европейско-сибирской группы – евро-байкальский D. marginatus и евро-обский A. subfuscus. Наиболее широко в Хибинах распространены бореальные и температные виды E. costalis, A. subfuscus, L. affinis и S. brunneus, фоновые в подстилках зональных сосняков и ельников (рис. 7). Общими для трех гор оказались лишь 3 вида щелкунов из 10: H. rivularius, E. costalis, L. affinis). Преобладание E. costalis в тундровых и лесотундровых биоценозах разных гор соответствует его характеристике как наиболее холодоустойчивого вида щелкунов, личинки которого имеют крайне низкие температуры переохлаждения: до -12 …-16 °С [Россолимо, 1994].
|
Рис. 7. Структура элатеридокомплексов в экосистемах Хибин. |
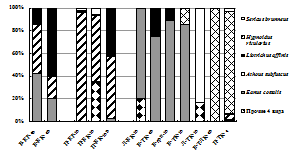
Проволочники большинства видов принадлежат к подстилочной экологической группе, их личиночное развитие связано с лесными местообитаниями. Это отражает «таежный» облик хибинской фауны щелкунов. По типу питания преобладают виды, которые сочетают сапро-, некрофагию и необлигатное хищничество и не повреждают живые растения. Имаго щелкунов, обитающих в Хибинах, ведут открытый образ жизни и активны в дневное время суток. Виды с сумеречно-ночной и смешанной активностью (самцы активны в течение суток, самки – только в сумерки или ночью) единичны в зональных биоценозах (роды Elater, Agriotes) и не встречены в горных.
С учетом наших данных элатеридофауна Хибин насчитывает не менее 10 видов из 10 родов, а Мурманской обл. в целом – 40 видов из 22 родов относящихся к 4 п/сем. при наибольшем видовом разнообразии родов Selatosomus и Ampedus (по 5 видов).
7.2. Дождевые черви (Oligochaeta: Lumbricidae)
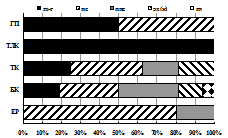
В биоценозах Хибин известны 4 вида дождевых червей (D. octaedra, D. rubudus, A. caliginosus и L. rubellus): от 3-4 видов с численностью до 130 экз./м2.в межгорных березняках до одного (D. octaedra) на вершинах гор [Рыбалов, 2006]. Нами выявлено 6 видов в основном почвенно-подстилочной жизненной формы: в горных ельниках 4, в лесотундровых березняках 5, в альпийских тундрах 6 видов (табл. 9).
Спектр видов и жизненных форм червей варьировал как по горам и склонам, так и по растительным поясам при наибольшем разнообразии в субальпийских березняках на южных склонах. Лиственным лесам с большими запасами подстилки, ее меньшей кислотностью и повышенным по сравнению с горными ельниками содержанием минеральных элементов, углерода и азота [Морозова и др., 1976; Почвообразование …, 1979] соответствовали высокие показатели численности и биомассы червей: до 55 экз./м2 и 8 г/м2 на г. Поачвумчорр, до 70 экз./м2 и 12 г/м2 на г. Вудъяврчорр. Обитание крупного теплолюбивого вида E. fetida в субальпийских березняках на склонах южной и юго-восточной экспозиции свидетельствует об интразональном характере этих биоценозов и полугидроморфном режиме горных почв.
Горно-лесные пояса Хибин (таежный и лесотундровый) характеризовались разнообразием морфо-экологических типов червей, которые образуют сукцессионный ряд: подстилочные → подстилочно-почвенные → почвенные. Он соответствует стадийности процесса деструкции растительного опада [Hendrix & Bohlen, 2002; Eisenhauer et al., 2007] и является показателем более глубокой степени биотрансформации органического вещества в почвенном ярусе горно-лесных биоценозов по сравнению с зональными подзолами.
Табл. 9. Обилие (%) дождевых червей в почвах высотных поясов Хибин
Гора | Вудъяврчорр | Поачвумчорр | Ловчорр | |||||||||
Пояс | ЕР | БК | ТК | ТЛК | ЕР | БК | БК | ТК | БК | ТК | ГП | |
Экспозиция | св | ю | юв | с | с | ю | ||||||
Питающиеся растительным опадом на поверхности почвы | ||||||||||||
Лесные подстилочные | ||||||||||||
Dendrobaena octaedra (Sav.) | – | 35 | 20 | – | 16 | 8 | 10 | 100 | 100 | – | – | |
Почвенно-подстилочные | ||||||||||||
Dendrodrilus rubidus (Sav.) | – | 8 | 20 | – | 84 | 16 | 80 | – | – | – | – | |
Lumbricus rubellus (Hoff.) | 50 | 57 | 20 | – | – | 20 | 8 | – | – | – | – | |
Eisenia fetida (Sav.) | – | – | – | – | – | 20 | 2 | – | – | – | – | |
Eisenia nordenskioldi (Eisen) | – | – | 20? | – | – | – | – | – | – | – | – | |
Питающиеся почвенным перегноем | ||||||||||||
Собственно-почвенные среднеярусные | ||||||||||||
Aporrectodea c. caliginosa (Sav.) | 50 | – | 20 | – | – | 36 | – | – | – | – | – | |
Всего | видов | 2 | 3 | 4+1? | 0 | 2 | 5 | 4 | 1 | 1 | 0 | 0 |
экземпляров | 2 | 40 | 5 | 0 | 58 | 30 | 186 | 3 | 3 | 0 | 0 | |
На северо-восточном склоне г. Вудъяврчорр максимальное в высотно-поясном ряду разнообразие люмбрицид (5 видов) выявлено весной в тундре кустарничковой на высоте 370 м, при этом все виды были представлены единичными особями. Азиатский по происхождению вид E. nordenekioldi найден только в этом биоценозе и указывается для Мурманской обл. впервые.
ВЫВОДЫ
1. Почвенная фауна Хибин представлена не менее чем 28 таксонами. В ельниках горной тайге выявлено от 20 до 24 таксонов, в лесотундровом поясе 9-24, в тундрах 8-22. По численности во всех поясах преобладают насекомые (11-85%) и черви (8-71%), главным образом, двукрылые (18-77%) и энхитреиды (до 70%) с короткими циклами развития в почве. Доля паукообразных и многоножек не превышает 17%, моллюсков – 2%. С высотой снижается доля тонкопокровных червей и моллюсков и возрастает обилие членистоногих (многоножек и паукообразных).
2. Почвенная фауна горно-лесных поясов сохраняет ядро доминантов, характерных для подзолов зональной северной тайги (двукрылые, жесткокрылые, пауки, многоножки, энхитреиды, люмбрициды), но обеднена насекомыми разных трофических уровней. Горные тундры, напротив, отличаются от зональной тундры высокой численностью и разнообразием почвенной фауны, сопоставимым с зональной тайгой. Это объясняется мягкими климатическими условиями альпийского пояса в связи с положением Хибин в подзоне северной тайги, а самих тундр – на небольших высотах. О мягкости горного климата свидетельствует «таежный облик» фауны жесткокрылых и проникновение лесных видов жуков в альпийский пояс.
3. В высоко гумусированных и обогащенных биогенными элементами почвах Хибин хорошо развит комплекс сапрофагов с преобладанием ключевых гумификаторов – дождевых червей и энхитреид, что свидетельствуют о большей функциональной активности сапроблока в горных экосистемах по сравнению с зональной тайгой. Показателем более глубокой степени биотрансформации органического вещества в почвенном ярусе горно-лесных биоценозов является широкий спектр видов и морфо-экологических типов червей, которые образуют сукцессионный ряд: подстилочные → подстилочно-почвенные → почвенные, соответствующий стадийности процесса деструкции опада. В субальпийских березняках южных склонов высокие показатели численности (до 500 экз./м2), биомассы (до 12 г/м2) и разнообразия сапрофагов, отличающие структуру высокопродуктивных сообществ, свидетельствуют об интразональном характере этих биоценозов.
4. Фактор поясности в большей степени влияет на сапро - и фитофагов, в меньшей – на хищников. Среди зоофагов перераспределяется доминирование по численности и биомассе между таксонами при сохранении их состава от 5-7 таксонов в горно-лесных поясах до 6-7 в тундрах. Сапрофильный комплекс в горных тундрах обедняется крупноразмерными дождевыми червями и моллюсками (3-4 таксона против 6-7 в горно-лесных поясах). В комплексе фитофагов с высотой сужается спектр таксонов, насекомые-обитатели напочвенной растительности сменяются почвенно-подстилочными видами щитовок и жуками – пилюльщиками, листоедами, долгоносиками. Для ряда групп (жесткокрылые, многоножки, дождевые черви) высотно-поясное распределение сходно с широтно-зональным распространением в пределах региона. Для массовых групп жуков (стафилинид, жужелиц, щелкунов) подтверждена смена таежных видов тундровыми при переходе от таежного пояса к субальпийскому и, далее, к альпийскому. Во всех поясах по составу и численности преобладают виды с лесной преференцией.
5. Влияние экспозиции склона проявляется в более высокой функциональной активности сапроблока и достоверно высокой численности сапрофагов (люмбрицид, энхитреид, моллюсков, элатерид) в почвах южных склонов по сравнению с зональной северной тайгой. Почвенная фауна склонов северной и северо-восточной экспозиции по составу таксонов и численности ближе к таковой в зональных подзолах.
6. Период активной трофической деятельности сапрофагов в горных почвах ограничен 3-4 месяцами. Наиболее уязвимы к воздействию природно-климатических условий Хибин крупноразмерные тонкопокровные дождевые черви, моллюски и аэробионтные насекомые, личинки которых развиваются в почве. Экстремальными горными факторами, влияющими на численность, разнообразие и сезонную активность фауны являются: крутизна склонов, нестабильный гидротермический режим, значительная скорость ветра, короткий период с положительными температурами воздуха.
7. В горных биоценозах выявлено 79 видов жесткокрылых: 47 видов стафилинид из 27 родов, 7 п/сем.; 22 вида жужелиц из 18 родов, 4 7 п/сем.; 10 видов щелкунов из 10 родов, 7 п/сем. Впервые для Хибин указаны 42, 11 и 6 видов из этих семейств, впервые для Мурманской области – 19, 4 и 1 вид. Региональный список стафилинид расширен до 187 видов из 64 родов, жужелиц – до 69 из 26 родов, щелкунов – до 40 видов из 22 родов. С учетом литературных данных хибинская фауна стафилинид, жужелиц и щелкунов теперь насчитывает 56, 30 и 10 видов соответственно. Типичными обитателями Хибин можно считать лишь 12 видов, наиболее широко распространенных в горных биоценозах: стафилинид Mycetoporus lepidus, M. monticola, Atheta brunneipennis, A. graminicola, Oxypoda annularis, жужелиц Patrobus assimilis, Calathus micropterus, Amara brunnea, щелкуны Eanus costalis, Liotrichus affinis, Athous subfuscus, Hypnoidus rivularius.
Список публикаций по теме исследования
Публикации в изданиях, рекомендованных ВАК РФ
, В., Похилько к почвенной фауне Хибинского горного массива на примере горы Вудъяврчорр // Вестник МГТУ, 2009. Т. 12. № 3. С. 516-524.
, В., Похилько распределение почвенной фауны Хибин // Почвоведение, 2011. №9. С. .
, В., , Середюк поясность и комплексы жесткокрылых в почвах Хибинского горного массива // Труды Карельского НЦ РАН, 2011. №2. С. 107-118.
Публикации в тематических сборниках и трудах конференций
, В. Беспозвоночные животные в почвах горно-растительных поясов Хибин (на примере г. Вудъяврчорр) // Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт / Матер. междун. конф. – Мурманск: МГПУ, 2009. С.101-105.
В., Зенкова беспозвоночных в подстилках равнинных и горного ельников Мурманской области // Продуктивность и устойчивость лесных почв / Матер. междун. конф. по лесному почвоведению. – Петрозаводск, 2009. С. 218-220.
В., Зенкова жуков-щелкунов (Сем. Elateridae) в равнинных и горных почвах северотаежной подзоны Мурманской области // Энтомологические исследования в Северной Азии / Матер. VIII межрегион. совещ. энтомологов Сибири и Д. Востока. – Новосибирск, 2010. С. 169-171.
В., Зенкова (Coleoptera: Stafilinidae, Carabidae, Elateridae) в почвах Хибин // Экологические функции лесных почв в естественных и нарушенных ландшафтах / Матер. IV Всерос. научн. конф. по лесному почвоведению. – Апатиты: Кольский НЦ РАН, 2011. Ч. 1. С. 44-48.
, В. Разнообразие беспозвоночных животных в почвах тундровой и лесной зоны Мурманской области / Матер. всерос. конф. к 100-летию . – Якутск, 2011. С. 104-110.
, Зенкова мезофауна Хибинского горного массива (Мурманская область) // Проблемы почвенной зоологии / Матер. XVI всерос. совещ. по почвенной зоологии. – Москва, 2011. С. 90-92.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
