


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ДУБРОВСКИЙ Николай Григорьевич
СТЕПНЫЕ И ЗАЛЕЖНЫЕ ФИТОСИСТЕМЫ ТУВЫ:
структурно-функциональная организация и оптимизация
03.00.05 – Ботаника
03.00.16 – Экология
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Улан-Удэ
2009
Работа выполнена на кафедре общей биологии Тывинского государственного университета и на кафедре ботаники Бурятского государственного университета
Научный консультант:
доктор биологических наук, профессор
Бимба-Цырен Батомункуевич Намзалов
Официальные оппоненты:
доктор биологических наук, профессор Татьяна Петровна Анцупова
доктор биологических наук, профессор Иван Икидиусович Илли
доктор биологических наук Вера Ивановна Власенко
Ведущее учреждение:
Алтайский государственный университет,
г. Барнаул
Предварительная дата защиты « 29 » мая 2009 г. в ______ часов на заседании диссертационного совета Д 212.022.03 в Бурятском государственном университете г. Улан-Удэ, ул. Смолина, 24а, Бурятский государственный университет
Факс: (30; E-mail: *****@***ru
E-mail: *****@***ru
С диссертацией можно ознакомиться в библиотеке Бурятского государственного университета
Автореферат разослан « ____ » ___________ 2009 г.
Ученый секретарь
диссертационного совета,
к. б.н.
ВВЕДЕНИЕ
Актуальность темы. Изучение растительного мира – этой сложнейшей материальной системы нашей планеты, во всем его многообразии и на всех известных уровнях организации относится к числу важнейших задач биологической науки.
За последнее время в изучении растительного покрова Сибири сделаны большие успехи. Однако на обширных просторах Сибири отдельные части ее все еще остаются мало исследованными, к числу таких регионов и относится Тува.
Исследование современного состояния степей как наиболее широко распространенного типа растительности, слагающей основу ландшафтов как в межгорных котловинах, так и высоко в горах особенно актуально в связи с тенденцией нарастания антропогенной трансформации степных экосистем. Причем, это связано не только с пастбищной деградацией растительности, но и большими масштабами распашки степей, которые в настоящее время на больших площадях находятся в залежном режиме.
Касаясь региональных особенностей демутационных процессов, следует отметить факт развития особой мелкобурьянистой стадии, предшествующей развитию крупного бурьяна, в условиях абаканских и тувинских степей (Голубинцева, 1930; Дымина, 1983; Куминова, 1985). С другой стороны, на залежах по фону каштановых почв в Бурятии наблюдаются случаи выпадения бурьянистой стадии (Быков и др., 2003). В связи с этим возникла необходимость специального изучения особенностей зарастания заброшенной пашни, видового состава растительности, стадий зацелинения и их длительности, возможности восстановления плодородия почвы при зарастании залежей в Центрально-Тувинской котловине Республики Тыва. В настоящее время площадь пашни, подвергшейся стихийной консервации в республике по официальным данным составляет 59 тыс. га. Этой малоизученной и актуальной проблеме посвящена настоящая диссертация. Кроме этого, познание структуры и динамики растительного покрова степей, продуктивности и многообразного влияния антропогенных факторов на степные пастбища при преимущественном овцеводческом направлении хозяйства Тувы представляет также и большой практический интерес.
Цель и задачи исследования. Целью работы является выявление флористического и фитоценотического разнообразия степных и залежных фитоценозов Тувы, а также их структурно-функциональных особенностей в связи процессами демутации растительности степных залежей.
Для достижения поставленной цели предусматривались следующие задачи:
1. Выявить состав флоры степей и степных залежей осуществить систематический, биоморфологический и эколого-географический анализ;
2. Сформулировать основные положения принятой классификации и на ее основе дать полную геоботаническую характеристику выделенным таксономическим категориям степной и залежной растительности;
3. Установить основные стадии в сукцессии залежной растительности с характеристикой структуры сообществ и динамики продуктивности надземной фитомассы;
4. Исследовать микробные сообщества в опадах залежных фитоценозов и определить сезонную динамику численности микроорганизмов – бактерий деструкторов
5. Определить скорость микробиологической деструкции целлюлозы и белка в исследуемых растительных сообществах, а также изучить характер деструкции в зависимости от абиотических факторов окружающей среды (температуры и влажности)
6. Оценить современное состояние степных и залежных экосистем и выработать подходы к эколого-адаптивной концепции в природопользовании в Тыве.
Материалы и методы исследования. В ходе выполнения данной работы были проведены полевые исследования с гг. Собран гербарный материал, включающий более 2300 гербарных листов, сделаны более 720 полных геоботанических описаний. Для изучения видового состава растений использовали: «Определитель растений Тувинской АССР», «Флора Сибири» и другие источники.
Сбор материала производился c использованием общепринятых методов геоботанических и флористических исследований. Проведены исследования степных и залежных фитоценозов кожуунов: Барун-Хемчикский, Улуг-Хемский, Дзун-Хемчикский, Кызылский, Тандынский, Эрзинский, Овюрский, Монгун-Тайгинский и Пий-Хемский. Названия видов сосудистых растений приведены по флористической сводке (1995). Географические группы выделены по (1980). Жизненные формы выявлены по классификации (1964), экологические группы – по (1960).
При классификации растительности использован доминантно-детерминантный подход, сочетающий критерии флористической и эколого-фитоценотической классификаций, обоснованной во флороценогенетической концепции (1947), (1979), (1994). При систематизации фитоценозов применена компьютерная программа TURBO (VEG) для первичной обработки геоботанических описаний.
Исследования по микробиологической деструкции растительных остатков проводились в гг. в Центрально-Тувинской котловине на пробных площадках, соответствующих основным стадиям зацелинения залежных фитоценозов. Продуктивность наземной фитомассы исследуемых растительных сообществ определяли методом укосов, в пяти повторностях (Родин,1968). Исследование скорости разложения растительного опада проводили методом механической изоляции (Перель, Карпачевский,1968). Общую биологическую активность растительного опада (протеазная и целлюлазная) изучали методом аппликации (Теппер и др.,1987). Учет численности микроорганизмов проводили методом предельных разведений. Аэробные и анаэробные целлюлозоразрушающие бактерии (ЦРБ) выращивали в жидкой среде Гетчинсона (Романенко, Кузнецов,1974) с добавлением фильтровальной бумаги. Учет численности аэробных и факультативно-анаэробных протеолитиков, амилолитиков и глюколитиков проводили в агаризованной среде Пфеннига (Phenning, 1965) с добавлением 1,5% пептона, крахмала и глюкозы соответственно.
Научная новизна. Впервые изучен видовой состав высших сосудистых растений степей и растительности залежной растительности региона исследования и в результате установлен список флоры степей Тувы. Последняя составлена 785 видами высших сосудистых растений (272 рода и 54 семейства). Проведен разносторонний детальный анализ флоры.
Проведена оригинальная классификация видового состава растительности залежей Тувы (110 видов сосудистых растений, 27 семейств и 80 родов) с установлением трех категорий флористического комплекса залежей (ФКЗ), а именно – типично залежных, переходных и инвазийных (или случайно залежных) типов ФКЗ. Долевые соотношения последних в растительности залежей имеют индикационное значение по особенностям их демутации. Выявлено все таксономическое разнообразие степных сообществ Тувы. Осуществлена эколого-фитоценотическая классификация степной растительности с характеристикой выявленных синтаксонов. Выделено 28 формаций, 13 групп формаций, относящиеся к 4 классам формаций (или подтипам) степной растительности.
Впервые для Центрально-Тувинской котловины Тывы проведено комплексное изучение залежной растительности, выявлены особенности демутации, изучена динамика продуктивности фитоценозов, а также численности микроорганизмов, участвующих в деструкции опада. Определен вклад различных физиологических групп бактерий в аэробную и анаэробную деструкцию органического вещества растительных остатков. Даны количественные показатели о скорости разложения опада в залежных фитосистемах Тувы, изучено влияние абиотических факторов (температуры и влажности) на темпы микробиологической деструкции.
Изучены особенности пастбищного использования растительности степных экосистем, закономерностей их антропогенной трансформации и процессов демутации на залежах, а также определены приоритеты эколого-адаптивного землепользования как перспективной для сухостепных ландшафтов межгорных котловин Тувы.
Практическая значимость исследования. В результате проведенных исследований, впервые наиболее полно выяснена типологическая структура степных пастбищ Тувы, их продуктивность, хозяйственное состояние. Полученные материалы по флоре и фитоценотическому разнообразию степей и залежей могут быть использованы в сравнительных экологических и фитогеографических исследованиях растительности гор юга Сибири и Центральной Азии. Содержащиеся в работе материалы могут послужить ценным источником при подготовке определителя региональной флоры, при флористическом и геоботаническом районировании, а также для экологического мониторинга и своевременного принятия мер по сохранению биологического разнообразия экосистем и выявления отрицательных воздействий на среду обитания растений и сообществ. Анализ результатов исследований залежных экосистем Тувы позволит обосновать новую концепцию экологического земледелия для аридных и континентальных районов Внутренней Азии. Изменение показателей почвенного плодородия при выведении пашни в залежь выявили перспективность данного приема для его восстановления.
В целом, основными результатами научно-исследовательской работы будут пользоваться другие ведомства по охране и использованию природных ресурсов Тувы, Тывинский государственный университет и другие образовательные учреждения. Собранные материалы пополнили гербарные фонды Бурятского и Тывинского госуниверситетов. Они могут использоваться при подготовке специалистов биологов и экологов в курсах систематики и экологии растений, геоботаники и биогеографии.
Защищаемые положения.
1. Флора степей Тувы, насчитывающий 785 вида высших сосудистых растений, отражает особенности буферного положения территории на стыке бореальных и пустынно-степных экосистем. При этом позиции горностепных южносибирско-монгольских и центральноазиатских видов ксерофитной экологии наиболее значительны.
2. Эколого-фитоценотическая классификация степной растительности с использованием доминантно-детерминантного подхода позволила выделить 28 формаций, 13 групп формаций и 4 класса формаций (подтипов). Однако, объективизация выделения синтаксонов может быть достигнута с использованием метода экологической ординации сообществ степей.
3. Анализ видового состава растительности залежей Тувы (294 видов сосудистых растений, 110 – только в Центрально-Тувинской котловине) с дифференциацией на три типа флористических комплекса залежей (ФКЗ), а именно – типично залежных, переходных и инвазийных (или случайно залежных) позволяет прогнозировать динамические тенденции в процессах демутации и могут служить индикаторами стадий зацелинения залежей.
4. Микробиологическая деструкция растительного опада является важнейшим процессом, определяющим биологический круговорот элементов в природе. При этом вклад различных физиологических групп бактерий в аэробную и анаэробную деструкцию органического вещества растительных остатков имеют региональные особенности на различных стадиях демутации. Это касается прежде всего скорости разложения опада в условиях сухостепных экосистем Тывы.
Апробация работы. Материалы исследования доложены и обсуждены на Всероссийской научно-практической конференции с международным участием «Опыт и традиции этнического природопользования в Байкальской Сибири» (г. Улан-Удэ, 2003), Всероссийской научной конференции с международным участием «Проблемы сохранения разнообразия растительного покрова Внутренней Азии» (г. Улан-Удэ, 2004), на VIII Международном Убсунурском симпозиуме «Убсунурская котловина как индикатор биосферных процессов в Центральной Азии» (г. Кызыл, 2004), на VII Международной конференции «Природные условия, история и культура Западной Монголии и сопредельных регионов» (г. Кызыл, 2005), Всероссийской конференции «Биоразнообразие и пространственная организация растительного мира Сибири, методы изучения и охрана» (г. Новосибирск, 2005), Международной научно-практической конференции «Проблемы ботаники Южной Сибири и Монголии» (г. Барнаул, 2002, 2006), Пятом международном симпозиуме «Степи Северной Азии» (г. Оренбург, 2006), Региональном научно-методологическом семинаре «Этническая экология и традиционное природопользование на рубеже веков» (г. Улан-Удэ, 2006), V научной конференции РАЕ с международным участием «Экология и рациональное природопользование», (Египет, г. Хургада 2007), Всероссийской конференции с международным участием «Синантропизация растений и животных» (г. Иркутск, 2007), Международной научной конференции «Мир кочевой цивилизации: история и современность» (г. Чита, 2007), Международной научно-практической конференции «Биоразнообразие и сохранение генофонда флоры, фауны и народонаселения Центрально-Азиатского региона» (г. Кызыл, 2007), заседаниях кафедры ботаники БГУ и кафедры общей биологии ТывГУ (2004, 2005, 2008).
Исследования были поддержаны грантом РФФИ «Разработка стратегии сохранения редких растений и фитоценозов Тувы» (, №), Грант РГНФ «Комплексное социально-экологическое исследование особенностей номадного природопользования аборигенных этносов горных районов Южной Сибири. (, № а), Правительства Республики Тыва (№ 54, 2005).
Публикации. По теме диссертации опубликовано 32 работы, в том числе 5 монографий и статей, из них 7 в рекомендованных ВАК изданиях.
Объем работы. Диссертационная работа изложена на 345 страницах. Состоит из введения, 7 глав, выводов, списка литературы и приложения. Материал представлен 19 рисунками (фотографии, схемы, диаграммы) и 20 таблицами. Список литературы насчитывает 312 наименований.
ГЛАВА I. История исследования растительности степей и залежей
Первые содержательные геоботанические и флористические сведения о растительном мире Тувы появились более ста лет назад в отчетах экспедиций, исследовавших Центральную Азию. Это путешествия ( и гг.), A. В. Адрианова (1881 и 1883 гг.), -Гржимайло ( гг.) и (1907 г.) Исследователи, совершившие специальные ботанические экспедиции – (1903) и (1914), X. Принтц (Printz, 1921).
Новый этап ботанических исследований Тувы относится к периоду установления Танну-Тувинской Народной Республики в 1921 г., и он связан с работами экспедиции ВАСХНИЛ, начиная с 1934 г., под руководством (1950). В 1928 г. работала Саянская ботаническая экспедиция Томского университета под руководством (Ревердатто, 1931).
Третий этап изучения растительного покрова, в частности, горных степей Тувы начинается с 1940-х гг. С 1945 по 1949 гг. исследованиями флоры и растительности Тувы занималась (1950, 1953, 1958). Кроме в е гг. ботанические исследования в Туве проводят (1953), (1957), (1964), (1975).
Четвертый, современный период ботанических исследований в Туве, начинается с 1970-х гг. Они начинаются с флористических работ и . Начиная с 1975 по 1981 гг., изучением растительного покрова Тувы занималась лаборатория геоботаники ЦСБС СО АН СССР, под руководством . Исследования степной растительности Хемчикской котловины в Западной Туве ведет (1978; 1979, 1982, 1985). Различным аспектам анализа структуры растительности и флоры горных степей региона посвятили свои работы и (1976, 1977), (1975, 1980), (1977), (1982а, б), (1982), (1982), (1981), (1982), и др., (1982), (1982), (1986) и др.
После некоторого перерыва, начиная с 1995 г. и, особенно с 2000 г. возобновляются исследования растительности степных экосистем региона с приоритетом анализа трансформации степей, изучения структурно-функциональных их особенностей. В постановке и реализации исследовательских программ степных экосистем велика роль Убсу-Нурского центра биосферных исследований под руководством и , а также естественнонаучных кафедр Тывинского госуниверситета с участием и координацией , , и . Итоги исследований последнего времени отражены в ряде диссертационных работ. Самбуу (2003,2004) и (2002), посвященны анализу продукционно-деструкционных процессов сухих степей Убсу-Нурской котловины. Рассмотрению геоэкологических аспектов функционирования степных ландшафтов песчаных массивов Тувы посвятила свои работы (2006). Степи нагорья Сангилен, особенности ее флористического состава, фитоценотического разнообразия и пространственной организации стали объектом исследования (2006). Флористические исследования в регионе продолжил (2007), раскрывший структуру флоры плато Алаш.
Таким образом, степень изученности степной растительности региона такова, что необходимо обобщение имеющихся материалов. При этом особого внимания заслуживают анализ результатов трансформации степей (пастбищного и залежного) и выявление их динамических тенденций. Залежная растительность Тувы до сих пор не была предметом специальных исследований.
ГЛАВА II. Природно-климатические особенности Тувы (орография, климат, почвы)
Тува, расположенная на юге Средней Сибири и примыкая к территории Монголии, приурочена между 49° и 52° с. ш. и 87° и 96° в. д., характеризуется сильно расчлененным рельефом. Современный макрорельеф Тувы сформировался на протяжении мезозоя и кайнозоя, причем орогенетическая активизация территории происходила по линиям древнейших тектонических разломов (Ефимцев, 1961; Девяткин, 1965; Чернов, 1988). В мезозое, в конце юры или раннего мела, устанавливается платформенный режим. В этот период на фоне медленного сводового поднятия территории преобладали процессы денудационного выравнивания. Речной сток из Тувинской котловины был направлен на юг, в сторону Гоби. Рельеф выглядел в виде низких гряд, холмисто-увалистых возвышений на фоне обширных выровненных равнин.
Новейший орогенный этап развития рельефа Алтае-Саянской области – этап альпийского тектогенеза вызвали блоковые поднятия, выявившие уже достаточно четко основные черты современного рельефа Тувы. Это оформление Саян, образование горстового хребта Западный Танну-Ола (Маслов, 1948, Зятькова, 1969). В это же время в результате слабых восходящих движений сформировался ряд внутригорных впадин в Тувинской котловине, обособившие Хемчикскую, Улуг-Хемскую и Кызыльскую впадины, поднявшимися низкогорными грядами - Адар-Таш, Берт-Дат. Горные сооружения, испытавшие постепенное поднятие, приобрели черты резко расчлененного рельефа (Западный Саян, Шапшальский). Отдельные хребты, испытавшие более резкое вздымание, сохранили остатки поверхности выравнивания на высотах м над ур. м. (Западный Танну-Ола, нагорье Сангилен). В пределах плато и нагорий характерна наибольшая дифференцированность поднятий, на фоне среднегорного сильно расчлененного рельефа возвышаются отдельные вершины до м (Сангилен, Алашское плато) (рис. 1).
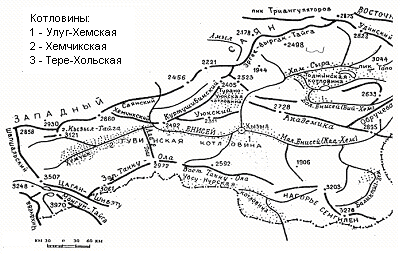
Рис. 1. Орографическая схема Тувы
Сложно организованная орография территории, располагающаяся в антициклоническом фасе крупнейших горных сооружений юга Сибири - Саян и Центрального Алтая, предопределила климатические условия Тувы. В целом территория находится в условиях умеренного и недостаточного увлажнения (Поликарпов и др., 1986). Распределе-ние осадков по региону чрезвычайно неравномерное. С одной стороны, территория недоступна воздействию влажных воздушных масс западного переноса; с другой, регион открыт мощным влияниям аридного и экстраконтинентального климата Центральной Азии. Наиболее экстремальный гидротермический режим создается в дождевой тени экранирующих хребтов (юго-западные части Хемчикской и Убсу-Нурской котловин, где сумма осадков достигает 80-150 мм в год). С другой стороны, создаются оптимальные условия на наиболее удаленных от магистральных хребтов, горных возвышениях, где транзитные воздушные потоки верхнего яруса тропосферы оставляют большую часть своей влаги. Такие ситуации создаются на северных макросклонах восточных отрогов Западного Танну-Ола, где выпадает до 700-мм осадков, тогда как на шлейфах южного макросклона этих хребтов – всего около 100-150 мм.
Сложный рельеф и контрастный климатический режим территории определяют большое разнообразие и пространственную неоднородность почвенного покрова Тувы. Для всех типов почв характерна малая мощность почвенного профиля из-за неглубокого проникновения почвообразовательных процессов в толщу материнских пород (Кононова, 1963). Почти все типы почв в горностепных ландшафтах отличаются грубым механическим составом, содержат большое количество щебнистого материала, на небольшой глубине подстилаются осадочными или кристаллическими материнскими горными породами (Зятькова, 1969).
Степные почвы тувинских котловин относятся к почвам каштанового ряда, представленных подтипами темно-каштановых, каштановых и светлокаштановых почв. На первом месте по распространению стоят каштановые почвы (Носин,1963), по механическому составу в основном легкосуглинистые и супесчаные разновидности с неглубоким (в пределах 50 см от поверхности) залеганием щебня или песка. В целом для каштановых почв характерны малая мощность, легкий механический состав, высокое содержание скелетных включений (до 50-80%), незначительные запасы гумуса - 2,3-4,0 %. Среди степных почв особого внимания заслуживают почвы верхних поясов аридных гор Тувы. Характерными чертами этих почв являются укороченный профиль, присутствие на поверхностях органно-генного грубогумусного горизонта с высоким содержанием в нем гумуса (до 12 %), слабокислой реакцией верхних горизонтов и щелочной - в нижних (Максимович, Ногина,1984).
ГЛАВА III. Методология изучения разнообразия фитосистем
Биологическое разнообразие как фундаментальное понятие классических наук о биоте (ботаника, зоология) включает все многообразие живых организмов растений, животных и микроорганизмов, а также их сообществ и ландшафтных комплексов. Одно из самых лаконичных определений понятия «биоразнообразие» (БР) дано «БР - разнообразие организмов и их природных сочетаний» (1992). Однако, автор принимая за базовый уровень видовое разнообразие по сути развивает α (альфа) - таксономическую концепцию БР, означающая разнообразие генофонда, начиная с популяционного уровня до фено - и генотипов.
За детализацию видового разнообразия на уровне особей и популяций, полагая что они отличаются по генетическим, фенотипическим и возрастным структурным особенностям, ратует (1996). Им обосновывается тезис о том, что «БР проявляется на всех уровнях организации живой материи от особей до биосферы» (с.31). Однако столь дробное подразделение категорий БР может привести к такому явлению как континуум БР. При этом мы можем «потерять» такую узловую точку проявления БР как вид. Вид, являясь системой популяций, включает все многообразие особей как структурную основу биологического вида и является удобной «инвентаризационной единицей» (Мэгарран, 1992).
Другая позиция в ориентации приоритетов к определению БР выработана в подходе (1992). По мнению автора ключевым является «разнообразие элементов пространственно-временной структуры биомов». Это прежде всего означает разнообразие экосистем как проявление ценофонда (Седельников,1997). Понятие «ценофонд» отражает разнообразие биохорологических структур от уровня элементарных экосистем (сообществ), их сукцессионных серий до биомов. Суть подхода - через выявление ценофонда, разнообразия зональных экосистем с учетом редких и исчезающих его компонентов, обеспечить сохранение потенциала БР видов и их генофонда. Этих же позиций придерживается (1994), по его мнению «зная структуру комплекса растительности, выражающуюся в наборе и площадном соотношении слагающих его растительных сообществ, мы можем оценивать и прогнозировать присутствие на исследуемой территории как групп, так и отдельных видов растений» (с.590). Данный подход в какой-то мере раскрывает β (бета) компоненту БР как разнообразие сообществ
Наиболее полным, с точки зрения принципов системности и иерархичности, остается подход Р. Уиттекера (1972). Понятие «Биоразнообразие» автор рассматривал как систему взаимодополняющих на разных уровнях структурных элементов экосистем (в данном случае экосистема рассматривается как надценотическая, ландшафтная единица). При этом уровень разнообразия видов (α), на следующем уровне рассматривается как ценотический (β) и наконец, более высокий уровень интеграции предусматривает БР определенных ландшафтов (γ/гамма/- разнообразие). Причем, разнообразие в пределах ландшафта означает многообразие как видов, так и сообществ (рис. 2).
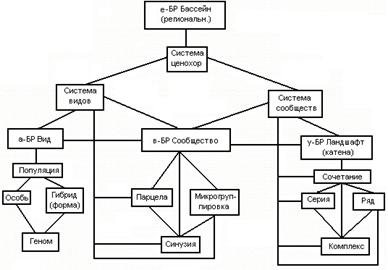
Рис. 2. Схема узловых уровней биоразнообразия фитосистемы
По мнению многих экологов видовое разнообразие намного сильнее связано с разнообразием сообществ и архитектурной сложностью местообитаний, т. е. ландшафтной структурой. Чем разнообразнее ландшафт, тем выше экологический объем местообитаний. Многообразие местообитаний по образному выражению (1981, 1986), является следствием "увеличения гиперпространства ниш сообщества" (1986, с.55). Последнее означает обеспечение полноты популяционного спектра видов, включая и временной фактор связанный с микроэволюционными процессами в сообществах (формирование видов). Выявление разнообразия сообществ как следствия реакций биоты на воздействия различных факторов среды и составляет цель измерения В-разнообразия. В данном случае ценоз или сообщество организмов является конкретным выражением β-разнообразия, также как особь - при определении ά-разнообразия. Разнообразие сообществ выражается числом таксонов или типов сообществ (ассоциаций, формаций, классов и т. д.).
Особый интерес вызывает следующий уровень БР (γ-разнообразие) биоты, разнообразие наименьшей биохорологической единицы - ландшафта. Данный уровень понимания БР до сих пор остается достаточно неразработанным. В частности (1992) считает целесообразным рассматривать β и γ - разнообразия как один тип БР. По мнению автора, смена сообществ по каким-либо градиентам (например, по ряду увлажнения) означает экологический диапазон изменений все того-же β-разнообразия, комбинации сообществ в границах ландшафтов. Подобные территориальные системы синтаксонов называются ценохорами (Сочава, 1972) или сигматаксонами (в подходах флористической класс-сификации растительности). Уиттекер (1972) γ-разнообразие рассматривал как общее разнообразие группы участков ά и β-разнообразий, но при этом не дал четких критериев: "что нужно считать базовой счетной единицей данного разнообразия?". Однако, Р. Уиттекер выделял особую категорию разнообразия (ε-эпсилон), как региональную. Сюда включались разнообразие группы территорий с однотипной генетической "конструкцией" ландшафтов. По сути это было разнообразие крупных биогеографических единиц (районов, провинций, областей).
После краткого рассмотрения общих теоретических позиций концепции биоразнообразия, рассмотрим схематическое представление методологии анализа биоразнообразия растительного мира (рис. 2). Уровень ά-разнообразия показывает таксономическое разнообразие видов, которые отражены в региональных флористических сводках, определителях растений.
β-разнообразие - разнообразие растительных сообществ или фитоценозов. Познание данного уровня начинается с установления группировок из сочетаний особей видов растений и систем их популяций в границах однородного контура земной поверхности минимальной размерности. Это исследования парцеллярной и синузиальной конструкции сообществ, явлений мозаичности и ярусности структуре сообществ, что крайне важно для определения механизмов их устойчивости. Пространственная разнообразие и временная динамика сообществ, выраженная в фенологических спектрах, сукцесссионных схемах и картографических моделях растительного покрова, а также типологическое разнообразие фитоценозов, представленная в виде иерархических схем классификации синтаксонов составляет содержание раскрытие B-разнообразия растительности.
Ландшафтное разнообразие растительности, которое трактуется как γ-разнообразие, остается еще малоразработанной. Пока не выработан единый подход в раскрытии данного уровня. В нашей трактовке это ландшафтно-геоботанические исследования, ориентированые на выявление ценотического разнообразия сообществ и внутриландшафтных структурных категорий в единстве с физико-географическими условиями - почвенными, геоморфологическими, климатическими, литологическими и т. д. Важнейшим является анализ пространственной структуры территориальных единиц растительности (ландшафтных комбинаций) с целью познания механизмов структурирования и интегрирования сообществ и комбинаций в единую "ткань" растительного покрова. Непременным условием работ является заложение эталонных полигонов с постановкой на них детального крупномасштабного картографирования растительности. Дальнейшая типизация (классификация) выделенных однородных (сообществ) и неоднородных (комбинаций) единиц растительности полигонов дает системное представление о структуре растительного покрова регионов.
В целом, из трех базовых уровней биоразнообразия (ά, β, γ) ключевым представляется ландшафтная, которая включает в себя элементы первых двух - конкретные особи видов растений и фитоценозы. Действительно, вид можно сохранить лишь сохранив сообщества, в котором он отмечается, а разнообразие ценозов является следствием разнообразия местообитаний, морфогенетических поверхностей рельефа. Отсюда каждая природная комбинация обеспечивает не только разнообразие ценозов, но и системы популяции видов растений, как структурных частей сообществ. В этом заключается системообразующая роль комбинаций ландшафтного уровня и поэтому, значимость исследований типов структур РП будет вызывать все больший интерес (Намзалов, Дубровский, 2007б). В настоящей работе в соответствии с изложенной выше концепцией биоразнообразия раскрыта видовая (ά) и ценотическая (β) организация растительности степей Тувы, включая их пастбищно - и залежно трансформированные варианты.
ГЛАВА IV. Степи Тувы, анализ структуры флоры степей и залежных земель
Степи в Туве являются широко распространенным типом растительности и занимают 2811 тыс. га. (Ершова, Намзалов, 1985). Как все островные степи Южной Сибири, они приурочены к межгорным понижениям и окаймляющими их со всех сторон предгорьями хребтов и нагорий. В Туве они слагают основу ландшафтов Тувинской (Хемчикская и Улуг-Хемская) и Убсунурской котловин, ряда периферийных небольших межгорных впадин (Эдыгейская, Карахольская, Турано-Уюкская) и встречаются по южным склонам в лесном поясе всех горных систем.
Основные степные регионы:
1. Тувинская котловина (Хемчикская, Улуг-Хемская и Турано-Уюкская впадины).
2. Убсунурский (северная окраина Убсунурской котловины и южный макросклон Западного и Восточного Танну-Ола).
3. Монгун-Тайгинский (горный массив Монгун-Тайга и южный макросклон хребта Цаган-Шибэту).
Флора степей Тувы включает 785 видов, принадлежащих к 272 родам и 54 семействам. Список флоры составлен на основании гербарных сборов автора, а также анализа публикаций (1953), с соавторами (1973, 1975, 1980), (1974, 1980), (1981), (2006), (2007) и других. Флора степей Тувы богата, что особенно наглядно видно, если сопоставить ее с флорами островных степей Алтая, Хакасии, Забайкалья. Согласно данным систематического анализа флоры, к многородовым семействам относятся: Asteraceae (31 род), Роасеае (25), Fabaceae (14), Rosaceae (10), Chenopodiaceae (13), Brassicaceae (23). По видовому составу все представленные семейства можно разделить на три основные группы:
1). В группу ведущих семейств, включающих от 20 видов и более, входят 13 семейств, их удельный вес в сложении флоры высокий – 83,7% (655 видов). Сюда входят следующие семейства: Asteraceae(120), Роасеае (89), Fabaceae (79), Rosaceae (42), Chenopodiaceae (37), Liliaceae (30).
2). Группа семейств, включающая от 4 до 19 видов, состоит из 13 семейств, в составе которых 80 видов (10,2% от флоры степей): Primulaceae (10), Umbelliferae (10), Polygonaceae (13), Caryophyllaceae (11), Gentianaceae (9), Campanulaceae (12) и др. Виды, объединенные в эти семейства, образуют группу разнотравья, ценотическая роль которой в настоящих крупнозлаковых и луговых степях становится одной из ведущих.
3). Группа семейств с небольшим числом видов (от 1 до 4) пред-ставительна: 25 семейств и 47 видов (6,1%). Многовидовыми родами являются: Artemisia (38), Astragalus (27), Potentilla (24), Allium (22) (табл.1). Для познания флоры тех или иных регионов не менее важны роды с небольшим числом видов, особенно монотипные. Во флоре степей Тувы это - Arctogeron gramineum, Cymbaria daurica, Nitraria sibirica, Nanophyton erinaceum и др. Эти виды имеют разный генезис и в комплексе хорошо отражают сложный процесс становления флоры исследуемого региона (Дубровский, Намзалов, Кыргыс, Самдан, 2007).
Таблица 1
Количество видов основных родов флоры степей Тувы и основных районов
Род | Количество видов | Число видов в районах | |||
шт. | % от общего состава | I | II | III | |
Stipa Festuca Carex Chenopodium Allium Potentilla Astragalus Oxytropis Saussurea Artemisia | 12 10 16 9 22 24 27 20 9 38 | 1,50 1,27 2,00 1,15 2,80 3,06 3,40 2,50 1,15 4,80 | 10 3 11 7 18 11 15 8 5 30 | 8 6 7 7 9 17 20 6 4 26 | 5 9 7 3 14 8 6 10 5 13 |
Общее число видов | 785 | 503 | 367 | 263 |
Примечание. I - Тувинская котловина, II - Убсунурский, III - Монгун-Тайгинский

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
