

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Лекция 4.(2ч.).
Тема: Закономерности наследования признаков при половом размножении.
1. Основные правила наследственности.
2. Законы Менделя.
3. Виды доминирования.
4.Летальное действие генов.
5. взаимодействие неаллельных генов.
6. Наследственность и среда.
1. Основные правила наследственности. Очень важный вклад в выяснение сущности наследственности внес Г. Мендель, опыты которого по скрещиванию растений лежат в основе большинства современных исследований. Применяя скрещивание растений, различающихся по контрастным признакам, Мендель в 1865 г. сформулировал идею о существовании наследственных факторов. Гибридологический метод, связанный с изучением характера наследования отдельных признаков и свойств, в значительной мере предопределил успех исследований Менделя и позволил ему выявить и сформулировать основные правила наследственности.
К основным особенностям гибридологического метода изучения наследственности относят:
1) использование в качестве исходных форм для скрещивания растений, отличающихся друг от друга сравнительно небольшим количеством (одна, две или три пары) контрастных признаков, и тщательный учет характера наследования каждого из них;
2) точный количественный учет гибридных растений, различающихся по отдельным признакам, в ряде последовательных поколений;
3) индивидуальный анализ потомства от каждого растения в ряде последовательных поколений;
4) недопустимость влияния чужеродного генетического материала на родительские расы и гибриды;
5) сохранение способности к размножению у гибридов и их потомков.
2. Законы Менделя. Для опытов по скрещиванию Мендель выбрал 22 сорта гороха, которые имели четкие альтернативные различия по ряду признаков: круглые или угловатые семена, желтые или зеленые семядоли, высокие или карликовые растения. Исследуя эти сорта методом самоопыления, Мендель убедился, что они представляют собой наследственно чистые формы. В ряде поколений они не давали никаких отклонений от стандартных признаков сорта. Мендель скрещивал растения, отличающиеся друг от друга по одному, двум или трем парам контрастных признаков. Часть гибридных растений Мендель скрещивал с исходными сортами, а остальные подверг самоопылению. Во втором поколении гибридов он внимательно изучал все выращенные растения и записывал результаты своих наблюдений. Полученные данные Мендель обработал математически и в результате выявил четкую закономерность передачи отдельных признаков от родительских форм к потомкам в ряде последовательных поколений.
Скрещивая растения, различающиеся между собой по одной паре альтернативных признаков (такое скрещивание называют моногибридным), Мендель обнаружил, что гибриды первого поколения проявляют только один признак из альтернативной пары. Так, при скрещивании сорта гороха, имеющего желтые семена, с сортом, имевшим зеленые семена, все гибридные семена были желтыми, вне зависимости от того, какой сорт использовали в качестве материнского. При скрещивании высоких растений с низкорослыми в первом поколении все растения были высокорослыми и т. д. На основании этих опытов Мендель пришел к заключению, что при скрещивании сортов, отличающихся по одной паре признаков, у гибридов первого поколения (F1) всегда имеется признак только одного из родителей. Такие признаки Мендель назвал доминантными (лат. dominantis — господствующий). Те признаки, которые у гибридов первого поколения не проявлялись, Мендель назвал рецессивными (лат. recessus — отступление).
Наблюдаемое явление Мендель назвал правилом доминирования, которое в настоящее время принято называть первым законом Менделя или законом единообразия гибридов первого поколения.
Подвергнув гибридные растения самоопылению, Мендель обнаружил, что среди потомков таких гибридов наряду с особями, несущими доминантный признак, появились особи с особенностями другого родителя, ранее не проявившимися. Отмеченный Менделем факт того, что рецессивный признак в скрытом виде проходит через поколение гибридов и вновь возникает в их потомках, привел ученого к идее о существовании ответственных за эти явления наследственных факторов. Для удобства обработки полученных данных Мендель применил буквенное обозначение наследственных факторов: для доминантного признака А, для рецессивного — а. Наследственно чистые доминантные особи получили обозначение АА, а чистые рецессивные особи — аа. Альтернативные наследственные факторы стали называть аллелями. Аллели можно охарактеризовать как различные (контрастные) формы одного гена, возникающие в результате мутации гена, либо как признаки, обусловленные одной парой аллельных генов. В дальнейшем организмы, имеющие пару одинаковых аллелей (АА или аа), стали называть гомозиготными, а особей с набором Аа — гетерозиготными.
Путем самоопыления растений первого поколения Мендель получил семена F2. Три четверти этих семян имели желтую окраску и одна четверть — зеленую. При выращивании и самоопылении растений из зеленых семян было обнаружено, что все эти растения образуют только зеленые семена.
Растения, полученные из желтых семян, при самоопылении проявили иные свойства. Из желтых семян F2 Мендель вырастил и подвергнул самоопылению определенное количество растений. Одна четверть этого количества образовала только желтые семена, в то время как три четверти растений образовали как желтые, так и зеленые семена в количествах, приближающихся к соотношению 3:1. Таким образом, растения третьего поколения разделились на две группы: группу растений, образовавшую только желтые семена, и группу растений, подобно гибридам F1 дававшую как желтые, так и зеленые семена в соотношении 3:1 (рис. 10). Неконстантные и константные растения в F3 были обнаружены в соотношении, близком 2:1.
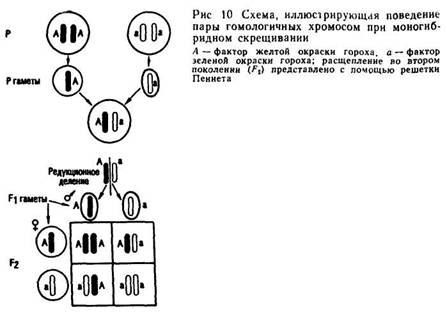
На основании проведенных опытов Мендель сформулировал второе правило, или закон расщепления признаков у гибридов второго поколения. Второй закон наследственности — основной вклад Грегора Менделя в экспериментальное изучение наследственности.
Третий закон Менделя — закон независимого наследования — был сформулирован им на основании изучения потомства от дигибридных скрещиваний. Так, при скрещивании сорта гороха, имевшего гладкие желтые семена (ААВВ), с сортом, имевшим зеленые морщинистые семена (aabb), гибриды F1 имели желтые гладкие семена, так как гладкая форма семян доминирует над морщинистой. В F2 среди 556 полученных семян
Мендель обнаружил 315 гладких желтых, 101 шероховатое желтое, 108 морщинистых зеленых и 32 морщинистых зеленых, что можно выразить соотношением 9:3:3:1. При рассмотрении этого соотношения видно, что наряду с семенами, обладающими сочетаниями признаков, характерными для исходных форм (гладкие желтые и морщинистые зеленые), в F2 появляются семена с новыми сочетаниями признаков — гладкие зеленые и морщинистые желтые, то есть происходит перекомбинация признаков исходных форм.
На основании подробного анализа данного опыта и ряда аналогичных скрещиваний Мендель пришел к заключению, что факторы А и В у гибридов АаВb при образовании половых клеток распределяются между собой совершенно независимо друг от друга и дают различные сочетания с одинаковой частотой. Поэтому среди половых клеток, образуемых гибридами F1, одна половина заключает фактор А, другая — а. Так же обстоит дело и с факторами В и b. Каждая половая клетка содержит только по одному представителю от пары факторов Аа и по одному представителю от пары Вb, поэтому возможны только четыре сочетания — АВ, Ab, aB, ab. Это в равной степени относится как к женским половым клеткам, так и к мужским.
Сочетания гамет с различными генотипами происходят совершенно случайно, поэтому образование зигот у гибридов F1 при дигибридных скрещиваниях математически можно выразить следующим образом: (AB + Ab + aB + ab) X (AB + Ab + aB + АВ). После раскрытия скобок и объединения зигот формула приобретает следующий вид: 9АВ : 3Аb : 3аВ : ab, который полностью соответствует эмпирически найденному соотношению: 9 гладких желтых (АВ) : 3 морщинистым желтым (Ab) : 3 гладким зеленым (аВ) : 1 морщинистому зеленому (ab).
Если предположить, что факторы А и В расположены в разных негомологичных хромосомах (независимое распределение наследственных факторов имеет место только при этом условии), то, пользуясь современной терминологией, весь процесс расщепления можно описать при помощи решетки Пеннета (рис. 11).
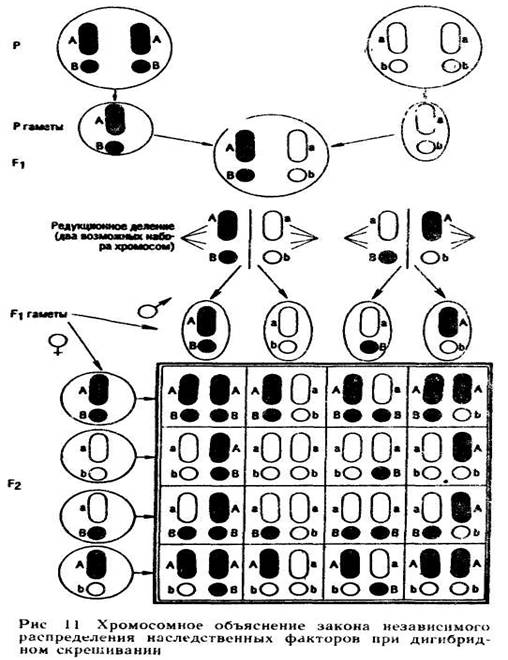
Для наглядности пара хромосом, в которых расположены гены А и а, изображена в виде палочковидных тел, а пара хромосом, в которых расположены гены В и b, — в виде округлых. Хромосомы доминантного предка окрашены в черный цвет, а хромосомы рецессивного оставлены белыми. Так как после мейоза число хромосом уменьшается вдвое, то гаметы доминантного родителя имеют генотип АВ, а гаметы рецессивного — генотип ab. Соединение гамет дает зиготу F1 с генотипом АаВb. Во время мейоза у гибридов F1 отцовские и материнские хромосомы расходятся в дочерние клетки совершенно независимо друг от друга, поэтому гаплоидные половые клетки с равной вероятностью могут содержать как две хромосомы одного из родителей (АВ или ab), так и по одной хромосоме от каждого из них (Аb или аВ).
Мужские и женские гаметы этих четырех групп соединяются между собой совершенно свободно, с равной вероятностью образуя зиготы, возникающие в результате сочетания мужских и женских гамет с любым из четырех генотипов: АВ, Аb, аВ и ab.
Решетка Пеннета, изображенная в нижней части рис. 11, дает представление о свободном сочетании гамет и генотипов зигот, возникающих в результате сочетания этих гамет. Зиготы, выписанные внутри решетки, образуют комбинационный ряд, состоящий из девяти членов, которые отличаются друг от друга по генотипу. Частоту встречаемости этих девяти членов можно записать следующим образом: AABB + AAbb + aaBB + aabb + 2AABb + 2aaBb + 2AaBB + Aabb + 4AaBb.
При условии полного доминирования члены, гетерозиготные по определенному фактору, внешне неотличимы от членов, гомозиготных по доминантному фактору, и объединяются с ними в одну фенотипическую группу. В данном случае выделены четыре фенотипических группы: АВ — гладкие желтые, Аb — гладкие зеленые, аВ — морщинистые желтые, ab — морщинистые зеленые.
После подсчета и объединения в группы фенотипически сходных зигот получается соотношение 9АВ : 3Аb : ЗаВ : lab, подобное соотношению, полученному Менделем.
В наиболее общей форме алгебраическое выражение расщепления при полигибридных скрещиваниях может быть представлено следующим образом:
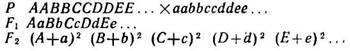
Установленные Менделем правила позволяли для каждого из более сложных случаев гибридизации определить, какое число особей охватывает общая формула, сколько в этой формуле членов и сколько членов будут гомозиготны, а также число сортов гамет (равное числу гомозиготных форм в, F2), образуемых гибридами F1. Согласно этим правилам, если число пар признаков, которыми отличаются исходные формы, равно п, то для F2 число особей в общей формуле равно 4п(4 у моногибридов, 16 у дигибридов, 64 у тригибридов, 256 у тетрагибридов и т. д.).
Число членов (число различных генотипов) равно 3п(3 у моногибридов, 9 у дигибридов, 27 у тригибридов, 81 у тетрагибридов и т. д.).
Число гомозиготных форм (и число фенотипов при полном доминировании) равно 2п (2 у моногибридов, 4 у дигибридов, 8 у тригибридов, 16 у тетрагибридов и т. д.).
Число сортов гамет равно 2п(2 у моногибридов, 4 у дигибридов, 8 у тригибридов, 16 у тетрагибридов и т. д.).
3. Виды доминирования. Соотношения в расщеплении у( гибридов второго поколения, установленные Менделем, наблюдаются при полном доминировании, то есть когда один аллель полностью подавляет действие другого. Впоследствии были обнаружены и другие виды доминирования. Например, при скрещивании безухих овец и баранов с нормально развитыми ушами получается короткоухое потомство. Такое явление называют неполным доминированием. При скрещивании коров с белыми пятнами на туловище, белыми брюхом и ногами (пегих) с быками, у которых окраска тела сплошная, получается потомство, имеющее белые пятна лишь на брюхе, голове, ногах, иногда большие пятна на туловище; следовательно, сплошная окраска неполно доминирует над пегой. К тому же у разных животных первого поколения размеры белых пятен колеблются от маленького белого пятнышка на вымени до почти сплошь белого брюха и т. п., следовательно, степень неполного доминирования у разных животных различна.
Впоследствии был обнаружен еще один тип доминирования — сверхдоминирование, при котором у гибридов первого поколения наблюдается более сильное развитие признака, чем у исходных родительских форм. Исследователи, открывшие это явление, объясняют его по-разному, однако большинство из них склоняются к тому, что при сверхдоминировании доминантный ген в одной дозе (то есть в гетерозиготном состоянии) более благоприятно влияет на развитие признака, чем в двойной. На явлении сверхдоминирования частично основан широко распространенный в США способ получения гибридных семян кукурузы путем комбинации при скрещивании четырех неродственных линий.
Еще задолго до открытия сверхдоминирования профессор высказал предположение о существовании таких генов, которые влияют на усиление развития признака, когда находятся в гетерозиготном состоянии, и, наоборот, могут ослаблять его и даже отрицательно влиять на организм — в гомозиготном. Такие гены предложил называть облигатно-гетерозиготными. Его гипотеза получила подтверждение при изучении гемоглобинов человека. Оказалось, что люди, гомозиготные по нормальному гемоглобину Л, заражаются тропической малярией и тяжело переносят эту болезнь; люди, гомозиготные по гену гемоглобина S, погибают из-за ненормальной формы эритроцитов (так называемой серповидно-клеточной анемии), а гетерозиготные не заболевают тропической малярией или легко ее переносят.
Не так давно установлен еще один вид доминирования — кодоминирование, при котором у гибридов первого поколения проявляются признаки того и другого предка, выраженные в равной степени и независимо друг от друга. Так, при скрещивании красных шортгорнских коров с белыми шортгорнски-ми быками получаются телята чалой масти (смесь белых и красных волос), и по фенотипу потомков легко определить, что они гетерозиготны по этим генам. По типу кодоминирования наследуются, в частности, группы крови животных и человека, различия в строении белков, например гемоглобина или транс-феррина, ферментов и пр.
4. Летальное действие генов. Французский зоолог Кено обнаружил, что при скрещивании между собой желтых мышей окраска их волосяного покрова никогда не закрепляется и в потомстве всегда дает расщепление на желтых и нежелтых в соотношении 2:1. Анализирующее скрещивание желтых и нежелтых особей давало расщепление 1:1. Результаты дальнейших опытов привели в конце концов к заключению, что все желтые мыши гетерозиготны, а зиготы, гомозиготные по желтой окраске, погибают на ранней стадии развития. Так впервые было показано, что ген в гомозиготном состоянии может быть летальным (смертоносным).
Среди овец каракульской породы встречаются животные с красивой серебристо-серой (ширази) окраской шкурки, оцениваемой дороже, чем обычный черный каракуль. При скрещивании серых овец с серыми баранами оказалось, что они всегда гетерозиготны, так как в их потомстве всегда было 25% черных ягнят. В то же время 25% ягнят серой окраски погибали от хронической тимпании при переходе на грубый корм. Причиной заболевания оказалось нарушение парасимпатической нервной системы. Когда же скрещивали серых баранов с черными матками или серых маток с черными баранами, то в потомстве оказывалось 50% серых и 50% черных ягнят, причем серые ягнята не заболевали. Оказалось, что в гомозиготном состоянии ген, вызывающий развитие серой окраски, обладал рецессивным летальным действием, основанным на нарушении функций парасимпатической нервной системы.
При искусственном осеменении, когда от одного быка можно получить несколько тысяч телят в год, опасность распространения вредных генов очень велика. Производитель, гетерозиготный по летальному гену, передает этот ген половине своих потомков, и если среди спаренных с ним коров не найдется достаточного числа гетерозигот по этому же гену, от которых может быть получено значительное число гомозиготных дефектных телят, то обнаружить носителя такого вредного гена можно только через несколько поколений.
Летальные гены, случалось, мигрировали из одного стада в другое и даже из одной страны в другую. Так, быка голштин-ской породы Принца Адольфа, завезенного из Голландии в Швецию в 1902 г., считали выдающимся производителем. Спустя 26 лет, когда стало известно, что он был носителем летального гена бесшерстности, его наследственный материал оказался широко распространенным среди шведских голштинов. Бык айрширской породы Данлоп-Талисман, завезенный в 1923 г. в Финляндию, обладал геном, вызывающим гидроцефалию. К концу 1937 г. в племенной книге айрширского скота Финляндии было зарегистрировано 84 сына и 189 внуков этого быка. Установили, что 82 его внука происходили от тех сыновей Данлоп-Талисмана, которые были гетерозиготами по данному летальному гену.
Сами по себе летальные гены не элиминируются; концентрация вредных генов в той или иной породе может достичь такого уровня, что возникает необходимость в специальных мерах для снижения частоты этих генов. Основной задачей при этом является выявление животных, в особенности производителей, гетерозиготных по нежелательному гену. Если такой ген полностью рецессивен, то выявить гетерозигот можно только с помощью испытания по потомству.
К настоящему времени у крупного рогатого скота определено 46 летальных генов, у лошадей 10, у свиней 18, у овец 15, у кур 45, у индеек 6, у уток 3. Различие в количестве леталей, обнаруженных у животных разных видов, зависит не столько от биологических особенностей вида, сколько от степени его изученности, количества и материальной ценности получаемого потомства, а следовательно, от большего или меньшего внимания к особенностям погибших животных.

Множественный аллелизм. До сих пор мы рассматривали только гены, существующие в двух аллельных состояниях, — доминантном и рецессивном, однако многие гены имеют более двух аллелей. Серией множественных аллелей называют три или более состояний одного локуса, обусловливающих разные фенотипы. Среди представителей одного вида аллели наследуются таким образом, что каждая особь может получить любые два из них, но никак не больше. В других сериях множественных аллелей гены могут быть не полностью доминантными, так что фенотип гетерозигот промежуточен между фенотипами родителей.
5. Взаимодействие неаллельных генов. Вышеизложенный материал содержит несколько упрощенное объяснение взаимодействия генов и признаков: каждый ген определяет один признак. Исследования показывают, что взаимодействие генов и признаков весьма сложное. Несколько пар генов могут определять появление одного признака; одна пара генов может извращать или подавлять эффект действия другой пары; определенный ген может вызывать различные эффекты в зависимости от изменения внешних условий. Гены наследуются как самостоятельные единицы, но они могут взаимодействовать сложным образом, определяя признак. Ниже приведены основные типы взаимодействия неаллельных генов.
Комплементарное действие генов. Примером одного из сравнительно простых типов взаимодействия может служить наследование формы гребня у кур. Известно, что ген розовидного гребня R доминирует над геном простого гребня r. Другая пара генов определяет различие между гороховидным Р и простым р гребнями. Следовательно, петух с простым гребнем должен иметь генотип ррrr, с гороховидным — РРrr или Ррrr, а петух с розовидным гребнем — ppRR или ppRr. Исследователи обнаружили, что при скрещивании гомозиготной птицы, имеющей гороховидный гребень, с гомозиготной, имеющей розовидный гребень, у потомков гребень оказывается не гороховидным и не розовидным, а совершенно иного типа, называемого ореховидным. Следовательно, ореховидность гребня определяется наличием у птицы одного или двух генов R и одного или двух генов Р, так что все куры генотипов PPRR, PpRR, ppRr и PpRr будут фенотипически одинаковы, имея ореховидный гребень. В результате скрещивания между собой двух гетерозиготных птиц с ореховидным гребнем в потомстве наблюдается расщепление в соотношении 9 ореховидных : 3 розовидных : 3 гороховидных: 1 простой (рис. 12).
Две независимые пары генов могут взаимодействовать таким образом, что ни один из доминантов не может вызвать соответствующего эффекта в отсутствие второго доминанта. Такие пары генов называют комплементарными (взаимодополняющими); в определении фенотипического эффекта действие каждого из них дополняет действие другого. Присутствие двух доминантов обусловливает появление одного признака, а противоположный признак возникает при отсутствии любого из них или обоих. Примером такого взаимодействия может служить образование коричневого пигмента у шелковичного червя.
Коричневый пигмент для насекомых имеет важное значение и связан с развитием фототаксиса. У тутового шелкопряда выявлены две рецессивные мутации двух неаллельных генов, обозначаемых W1 и W2; обе приводят к полной потере коричневого пигмента. Гибриды первого поколения от скрещивания двух форм, лишенных коричневого пигмента, из которых одна гомозиготна по гену w1(wwWW), а другая по гену w2 (WW w2 w2), имеют коричневый пигмент, так как содержат гены W1 и W2 (W1 w1 W2 w2).
В F2 идет расщепление на формы, имеющие коричневый пигмент, и формы, лишенные этого пигмента, в соотношении 9 : 7. Установлено, что образование коричневого пигмента связано с кинуренином и оксикинуренином, для которых, в свою очередь, исходным продуктом является триптофан. Вместе с пищей (листья шелковицы) тутовый шелкопряд в изобилии получает триптофан, который под влиянием соответствующих ферментов претерпевает ряд химических изменений и превращается сначала в кинуренин, затем в оксикинуренин и, наконец, в коричневый пигмент.
Ген w1 (в гомозиготном состоянии) вызывает образование белка, не имеющего ферментативных свойств, определяющих превращение кинуренина в оксикинуренин. Животные, гомозиготные по гену w1, лишены оксикинуренина, и у них накапливается значительное количество кинуренина Ген w2 в гомозиготном состоянии обусловливает отсутствие фермента, контролирующего превращение оксикинуренина в коричневый пигмент, поэтому животные, гомозиготные по гену w2, также лишены коричневого пигмента, но у них накапливается не кинуренин, а оксикинуренин.

Гибриды F1 от скрещивания генотипически различных форм, лишенных коричневого пигмента (W1 w1), гетерозиготны по генам W1 и W2. Имея в одинарном количестве нормальные аллели генов w1 w2, они способны превращать кинуренин в оксикинуренин и оксикинуренин в коричневый пигмент.
Во втором поколении некоторые животные несут ген w1 или w2 в гомозиготном состоянии и лишены пигмента.
Ранее было отмечено, что лишенные коричневого пигмента исходные формы w1 w1W2W2 и W1W1 w2 w2 отличаются друг от друга по содержанию кинуренина и оксикинуренина, поэтому в F2 среди особей без коричневого пигмента следовало ожидать 4/7, накапливающих кинуренин, и 3/7, накапливающих оксикинуренин. Действительно, среди особей F2 примерно в равном количестве были найдены особи, лишенные коричневого пигмента и содержавшие оксикинуренин, и особи, лишенные коричневого пигмента и содержавшие кинуренин Таким образом, не только установлен характер расщепления в F2, обусловленный взаимодействием двух неаллельных генов, но и выявлены биохимические процессы, которые обусловлены этим взаимодействием.
Эпистатическое действие генов. Как известно, доминирование есть подавление одним аллелем действия другого: А>а, В>b, С>с и т. д. Но существует взаимодействие, при котором аллель одного из генов подавляет действие аллелей других генов, например, А>В или В>А, а>В или b>А и т. д. Такое явление называют эпистазом.
Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию.
Гены, подавляющие действие других генов, называют супрессорами. Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных и у растений.
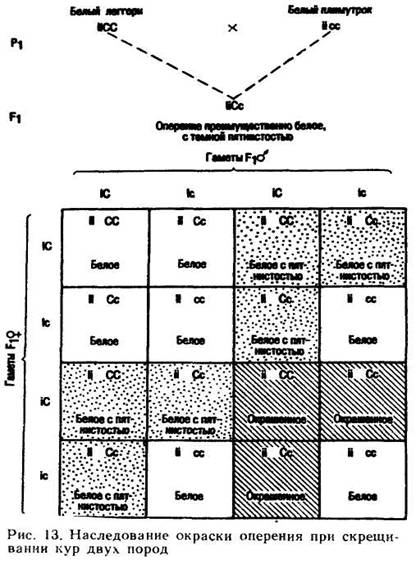
Изменение расщепления при таком типе взаимодействия генов можно наблюдать на примере скрещивания кур доминантной и рецессивной белой окраски. Белое оперение виандотов, плимутроков и некоторых других пород обусловлено отсутствием гена С, определяющего образование в перьях пигмента меланина. Перечисленные породы не являются полными альбиносами, так как глаза их окрашены. В отношении окраски оперения их генотип — ее окраска оперения леггорнов такая же, как и у других белых пород, но генетический анализ указывает на то, что они несут ген С. Действие этого гена подавляется у белых леггорнов геном-супрессором I, который в гомозиготном состоянии препятствует проявлению окраски. О белых леггорнах говорят, как о доминантных белых, потому что при скрещивании с окрашенными породами они дают в F1, преимущественно белое потомство. При скрещивании белых плимутроков и белых виандотов с окрашенными породами в F1 наоборот, получают исключительно окрашенное потомство, поэтому эти породы считают рецессивными белыми.
Когда белых леггорнов скрещивают с рецессивными белыми породами, то F1 обычно не чисто-белое, так как ген I у гетерозигот не полностью эпистатичен по отношению к гену С. У большинства цыплят и взрослых особей генотипа И встречаются черные пятна на пере, но в основном у них преобладает белое оперение; некоторые особи бывают совершенно белые. При скрещивании между собой гибридов F1 в F2 (рис. 13) в результате взаимодействия генов I и С и их аллелей расщепление по окраске будет следующим: 13 белых: 3 окрашенных. Если бы можно было идентифицировать каждую особь, гетерозиготную по I, то соотношение изменилось бы таким образом: 7 (чисто-белых) : 6 (белых с черными пятнышками) : 3 (окрашенных). Среди семи чисто белых три должны быть гомозиготными только по гену доминантной белой окраски, три —только по гену рецессивной окраски, а одна — по генам обеих белых окрасок, однако распознать эти три генотипа можно лишь путем анализирующих скрещиваний.
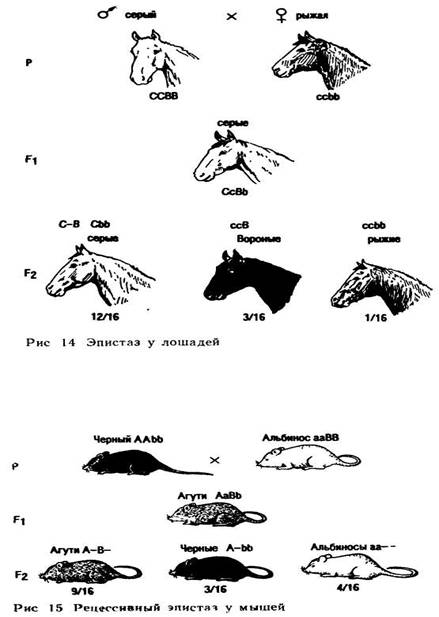
Подобную закономерность можно наблюдать при изучении наследования масти у лошадей. Ген С определяет серую масть, его рецессивный аллель с — рыжую. В другой паре хромосом имеется доминантный ген В, определяющий вороную масть, его рецессивный аллель b — рыжую масть При скрещивании серых лошадей с генотипом ССВВ и рыжих с генотипом ccbb в F1 получим гетерозигот СсВb серой масти, так как ген С эпистатичен гену В. Во втором поколении будет наблюдаться расщепление: 12 серых: 3 вороных: 1 рыжая. Данное соотношение свидетельствует также о расщеплении по двум генам: его можно представить как 9(С—В—)+3(С—bb) + 3(ccВ—) +1 (ccbb), что соответствует формуле 9:3:3:1. Очевидно, что в этом случае все лошади, имеющие ген С, будут серой масти. Лошади которые имеют ген В, но не имеют гена С, будут вороными и гомозиготное рецессивы будут рыжей масти (рис. 14).
В некоторых случаях супрессором можег быть и рецессивный ген. Такое явление называют «рецессивный эпистаз». Так, у мышей имеются ген А, определяющий серую окраску шерсти, и рецессивный ген а (черная окраска). В другой паре аллелей доминантный ген В способствует образованию пигмента, а его рецессивный аллель b подавляет синтез пигмента и мыши становятся альбиносами. Таким образом, гетерозиготные по обоим генам мыши (АаВb) имеют серую окраску. При скрещивании их между собой получится расщепление в соотношении 9 серых: 4 белых : 3 черных (рис. 15).
Полимерное взаимодействие генов. Это наиболее важная форма взаимодействия неаллельных генов, так как с ней связано выяснение такого важного вопроса, как наследование количественных признаков Полимерией называют такой тип взаимодействия генов, при котором на один признак действуют несколько пар неаллельных генов.
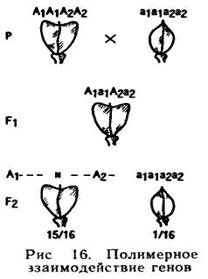
Например, у растения пастушья сумка известны две разновидности, одна из которых имеет плоды (стручки) треугольной формы, а другая — овальной. Гибриды F1, полученные от скрещивания этих разновидностей, имеют треугольные плоды. В F2 идет расщепление по форме плодов в соотношении: 15 растений с треугольными плодами : 1 растение с овальными. Если обозначить гены, принимающие участие в определении формы плодов, как А и В, и предположить, что для того, чтобы образовались плоды треугольной
формы, достаточно хотя бы одного из доминантных генов — А или В, то в результате такого скрещивания в F2 будет расщепление в соотношении 15:1. Растения, в генотипе которых имеется хотя бы один доминантный ген — А или В, имеют треугольные плоды, и только растения с генотипом aabb образуют овальные, и вследствие этого обычное дигибридное расщепление в соотношении 9:3:3:1 меняется на 15:1 (рис. 16).
У пшеницы известны два типа окраски зерен: белозерные формы, лишенные пигмента в оболочке зерна, и краснозерные, содержащие в оболочке зерна красный пигмент. Красная окраска доминирует над белым цветом зерен. При скрещивании сортов пшеницы, имевших красную окраску зерен, с белозерным сортом в F1 зерна всех растений были промежуточной розовой окраски, а в F2 у 1/16 растений — красной, у 1/16 — белой, а у остальных 14/16 — промежуточной окраски разной степени интенсивности. Было отмечено, что интенсивность пигментации усиливалась с увеличением числа генов, влияющих на развитие пигмента. В данном случае происходит накопление действия генов, усиливающих развитие признака, когда действие их как бы суммируется и увеличение развития признака зависит от эффекта действия каждого из них. Такое действие генов называют аддитивным (суммирующимся), а сами гены, усиливающие развитие признака, — аддитивными. Следовательно, при аддитивном действии генов величина признака зависит от числа генов, влияющих на усиление его развития.
Модифицирующее действие генов. Генами-модификаторами называют специфический тип генов, ослабляющих или усиливающих действие основного гена, влияющего на развитие определенного признака. Примером действия генов-модификаторов может служить вариация белой пятнистости у айрширов или голштино-фризов. Обе породы гомозиготны по гену, определяющему возникновение белой пятнистости, но гены-модификаторы вызывают вариацию проявления пятнистости от почти полной пигментации всего тела до почти полного ее отсутствия.
6. Наследственность и среда. Весь процесс развития особи, от оплодотворенной яйцеклетки до взрослого организма, происходит под непрерывным регулирующим влиянием генотипа, а также множества различных условий среды, в которой находится растущий организм. Свойства особи зависят, следовательно, от двух основных факторов — генотипа и среды.
Внешние различия, которые зависят только от влияний среды, называют модификациями. При недостатке кормов коровы будут давать меньше молока, чем коровы, получающие обильные корма. Данный сорт пшеницы развивается гораздо лучше, если она получает достаточно азота, чем в тех случаях, когда количество азота ниже нормы. Типичные примеры модификаций, обусловленных чисто внешними влияниями, можно наблюдать на любом засеянном злаками поле. Следовательно, необычайная изменчивость, свойственная живым существам, в значительной степени вызвана тем, что разные особи почти всегда находятся в разных условиях среды. Однако не менее важна изменчивость, зависящая от генотипических различий, то есть от различий в генетической конституции. В первую очередь эти различия связаны с рекомбинацией генов.
Имеется ряд возможностей для получения особей, идентичных по своему генотипу, так, с помощью вегетативного размножения из одного растения можно получить большое количество одинаковых потомков в виде клона. В таком случае все особи внутри клона будут иметь один и тот же генотип. При выращивании растений одного клона в разных экологических условиях было получено большое количество модификаций, однако после возвращения растений в прежнюю среду обитания все модификационные свойства исчезают; они не проявляются и в последующих поколениях
Большой интерес представляет изучение особей, идентичных по генотипу. Такое явление наблюдается при рождении однояйцовых близнецов. Среди близнецов наиболее часто встречаются так называемые дизиготные, или неидентичные (разнояйцовые), близнецы. Они возникают при оплодотворении двух независимых яйцеклеток двумя независимыми спермиями. Эти близнецы ничем не отличаются от обычных, в разное время рождающихся братьев и сестер — неблизнецов. При рождении однояйцовых близнецов одна оплодотворенная яйцеклетка делится на два бластомера, которые, разъединяясь, дают начало двум независимым друг от друга эмбрионам. Генотипы таких близнецов тождественны.
Исследования идентичных близнецов много дали для познания генетики животных и человека Они позволили изучить вопросы о соотношении среды и генотипа в развитии особи. Задача исследователей состоит в установлении степени сходства (конкордантности) или несходства (дисконкордантности) в парах близнецов.
Установлено, что удельный вес генетической информации в развитии особи очень велик. Близнецы поражают своим физическим сходством, но по-разному зависят от влияния условий внешней среды Возникновение у животного инфекционного заболевания определяется средой, однако даже в отношении таких свойств, которые, казалось бы, вызываются чисто внешними воздействиями, нельзя пренебрегать ролью генотипа. В проявлении таких заболеваний, как грипп, рахит, туберкулез, роль генотипа вполне очевидна. Степень наследственной предрасположенности играет роль при любом заболевании.
Классическим примером, иллюстрирующим сущность учения о норме реакции, являются опыты с так называемыми гималайскими кроликами. У этой породы кроликов глаза лишены пигмента и поэтому выглядят красными, тело покрыто белыми волосами и лишь ноги, уши и хвост пигментированы. Эти особенности строго передаются по наследству, будучи детерминированы аллелем сh, который вызывает менделевское расщепление при скрещивании, так что, казалось бы, перед нами — наследование признака в виде определенного рисунка пигментов на теле кролика. Однако оказалось, что появление этого признака требует строгого соотношения генотипа с определенными условиями среды. Генотип кролика таков, что пигмент в клетке начинает вырабатываться лишь при пониженных температурах, поэтому на всех участках тела кролика, где кровоснабжение ухудшено, образуется черный пигмент. Если же выщипать волосы из участка белой шкурки кролика и позволить им вырасти вновь, но уже в условиях холода, то они вместо белых окажутся черными. Если выщипать волосы с пигментированных частей тела кролика и позволить им вырасти в условиях повышенной температуры, то вновь выросшие волосы окажутся белыми. Таким образом, наследуется не рисунок гималайского кролика как таковой, а способность в зависимости от температурных условий к образованию пигмента.
Способность организмов реагировать на факторы среды, как правило, имеет приспособительный характер, и это составляет важнейшую сторону учения о норме реакции. Способность противостоять колеблющимся условиям среды путем адаптивного реагирования организмов получила название физиологического гомеостаза. У человека при жизни на разных высотах над уровнем моря вырабатывается разное количество эритроцитов; их концентрация в 1 мм3 у людей, живущих на уровне моря, в два раза меньше, чем у людей, живущих высоко в горах. Переселение в места, расположенные на высоте уровня моря, приводит к уменьшению числа эритроцитов в крови.
Все перечисленное указывает на глубокие различия между признаком особи и геном. Признаки как таковые не наследуются, они развиваются лишь на основе взаимодействия генотипа со средой. Наследуется только комплекс генов, который определяет норму реакции организма, изменяющую проявление и выражение признаков в разных условиях среды.
Контрольные вопросы:
1. Что означают следующие термины: доминирование, рецессивность, аллель, генотип, фенотип, гомозиготность, гетерозиготность, кодоминирование, летальные гены, эпистаз, полимерия?
2. В чем суть открытий Менделя?
3. Что такое аддитивная полимерия?
4. В чем различие между доминированием и эпистазом? Приведите примеры действия генов-модификаторов.
