
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
25% (1988 г.) содержания натрия (p<0,05) с последующим его снижением (8,5-9,4%), и подъём уровня кальция в 1,5-2,2 раза от величин 70-х годов (Наточин и др., 1975, 1995). В это же период отмечаются существенные изменения в морфологическом составе крови (табл. 2). При этом отмечалась нестабильность
Таблица 2
Лейкоцитарная формула крови русского осетра (%)
Годы | Миелобл. | Нейтрофилы | Эоз. | Мон. | Лимф. | ИСЛ | ||
юные | зрелые | пат. | ||||||
1988 | 28,6 | 16,5 | 8,8 | невыд. | 23,6 | - | 22,5 | 3,4 |
1989 | 19,5 | 8,6 | 6,1 | 1,8 | 26,2 | - | 37,8 | 1,64 |
1990 | - | 9,0 | 6,5 | - | 20,6 | - | 63,9 | 0,56 |
Норма* | - | 1,6 | 5,0 | - | 12,8 | - | 80,6 | 0,24 |
Примечание: Миелобл. – миелобласты, пат. – патологические, Эоз – эозинофилы, Лимф. – лимфоциты, ИСЛ – индекс сдвига лейкоцитов. * Норма: Сухопарова, Сухенко, 1986
функционирования интерреналовой железы: вначале повышение активности, с интенсивным синтезом кортизола (12,15±1,8 нг/мл), затем - напряжение и истощение, при снижении поступления в кровь кортизола до 2,3±0,7 нг/мл (Баранникова и др., 1989, 1990; Баранникова, 1992). В это время уменьшается секреция в кровь половых стероидных гормонов примерно в 3 раза (1989 г.). Выявленное снижение биохимических и гематологических параметров крови в дальнейшем (1990 г.) сменилось тенденцией к нормализации, однако в отношении интерреналовой железы и кортизола такого не отмечалось.
Сходные изменения были зарегистрированы и у других видов осетровых - севрюги и белуги, но с присущими для этих видов особенностями.
Изменения в физиологическом состоянии осетровых в море отразились и на производителях, мигрирующих на нерест. Они проявились в усилении имевшихся изменений в биохимических показателях и появлением новых, ранее не встречавшихся. Практически все исследуемые показатели претерпели изменения в ту или иную сторону, в сравнении с 60-70-ми годами прошлого века, что можно видеть на примере самок русского осетра (рис. 1).
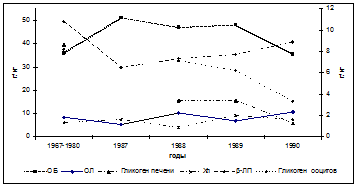
Рис. 1. Межгодовая изменчивость физиолого-биохимических показателей
у самок русского осетра
Также как и в море, у русского осетра, выловленного в дельте и под плотиной Волгоградской ГЭС, обнаружены аналогичные изменения в ионном составе крови: увеличение содержания кальция более чем в два раза у рыб, выловленных в дельте и в 1,6 раза под плотиной Волгоградской ГЭС (Кузьмина и др., 1992). Изменения в обмене веществ и минерального обмена сопровождались существенными колебаниями концентраций в крови кортикостероидных гормонов (Баранникова и др., 1989, 1990; Баранникова, 1992). Наиболее высокие концентрации в крови кортизола отмечались в весенние месяцы 1988 г., как у самок (48-40 нг/мл), так и у самцов (78-57 нг/мл) со снижением к июню до 16 нм/мл у тех и других. В дальнейшем имеет место относительная стабилизация его уровня с колебаниями в пределах 15-29 нг/мл. Примерно такая же динамика зафиксирована и в отношении половых гормонов прогестерона и тестостерона у самок и самцов. Похожая динамика отмечена и у эстрадиола. Негативным образом отразилось загрязнение среды обитания и на гематологических показателях производителей русского осетра, мигрирующего на нерест. Характер и направленность зафиксированных изменений (p<0,01) были схожими с таковыми в морской период жизни этих рыб с некоторым усилением. Наиболее рельефно они смотрелись у ослабленных производителей, подошедших к плотине Волгоградской ГЭС: СОЭ возрастало в 2 раза, при снижении в крови количества эритроцитов на 30%, в 1,6 раза лейкоцитов и 1,4 содержания гемоглобина, с трансформацией лейкоцитарной формулы. При этом индекс сдвига лейкоцитов возрастал в 4,6-5 раз от величин отмечаемых ранее. У самцов изменения гематологических параметров крови практически не отличались от самок. Не остаются неизменными в этих условиях и функциональные свойства гемоглобина: сродство гемоглобина к кислороду увеличивается в 1,3, а величина эффекта Бора 3,3 раза (Камшилов, 1990). В красной крови, наряду с увеличением гемолиза эритроцитов, наблюдается усиление кроветворения. Пониженная устойчивость эритроцитов к гемолизу, подтверждается и данными по осмотической резистентности эритроцитов (ОРЭ) крови русского осетра (Васильев, 1990). Она в 1988 и 1989 гг. была в 1,7-1,6 раза меньше, чем в 60-х годах.
У севрюги и белуги развитие реакции на загрязнение водной среды принципиально не отличалось от таковой осетра, хотя имели свои особенности.
Трансформации в обмене веществ, гормональном статусе и в целом в физиологическом состоянии повлияли на состояние половых желез осетровых рыб, в которых, выявленные в конце 80-х годов изменения, были настолько существенны, что привели к нарушению половой функции (Гераскин и др., 2001). Это не только большое разнообразие выявляемых патологий с появлением качественно новых форм, но и достаточно высокая частота встречаемости таких рыб. В море у рыб II СЗГ были обнаружены: гермафродитизм, интерсексуальность, опухоли, кисты и другие новообразования, отмечались сильные воспалительные процессы. В половых клетках изменения выявлялись, начиная с ооцитов протоплазматического роста. При переходе к вителлогенезу (II-III СЗГ) возрастает как частота встречаемости рыб с патологиями гамето - и гонадогенеза, так их разнообразие и степень.
Кроме того, морфофункциональные нарушения встречались в печени, почках, селезенке, сердце, скелетных мышцах. Наиболее значительными они были в неблагоприятные, с экологических позиций, годы (). В печени отмечено повышенное содержание липидов и липофусцина, белковая и жировая дистрофия, нарушения пигментного обмена, сосудистые патологии, хроническая воспалительная инфильтрация паренхимы, некробиоз гепатоцитов, цирротические явления. А в структуре её ткани обнаружены включения: мышц, половых желёз и ооцитов, селезёнки и даже участки с разными типами (до 4) тканей. В почках имело место явление белковой дистрофии и хронического межуточного нефрита, в селезенке – в основном сосудистые нарушения. В сердце нарушения проявлялись изменениями в волокнах и гиперемией кровеносных сосудов, имелись также очаги некроза (Евгеньева, 2000). Наиболее явный характер носили изменения в поперечнополосатой мышечной ткани (ППТМ). Выявлялись участки с нормальным строением ППТМ, слабо, умеренно и сильно выраженными изменениями этой ткани. На последних этапах развития этого процесса зафиксировано также снижение активности АТФ-азы вплоть до полного исчезновения. При далеко зашедшем патологическом процессе в ультраструктуре мышц наблюдалось снижение количества рибосом, исчезновение структуры аппарата Гольджи, полная деструкция митохондрий, фрагментация и набухание саркоплазматического ретикулума, изменение формы ядра (Басурманова, 1994). Конечной стадией разрушения мышечного волокна был полный лизис миофибрилл внутри волокна и разрушение сарколеммы. Впоследствии некротизированная мышечная ткань замещалась жировой или соединительной.
Морфологические изменения в мышцах сопровождались трансформацией их белкового состава проявлявшегося снижением количества выявляемых компонентов и изменением соотношения белковых фракций с большой, средней и низкой молекулярной массой (табл. 3). Наиболее существенные
Таблица 3
Фракционный состав водорастворимых белков белых мышц русского осетра (морской период жизни) с разной степенью морфологических нарушений
Уровень патологии (баллы) | Количество компонентов и их относительное содержание в белковых фракциях | |||
ФНМБ | ФСМБ | ФВМБ | ∑ | |
1-2 | 4 (2-5) 7,5±0,8% | 6 (4-7) 59,5±0,9% | 7 (5-8) 33,0±1,1% | 17 (12-19) 100,0% |
3 | 2 (1-4) 3,9±0,7% | 4 (2-7) 63,9±1,6% | 6 (5-7) 32,2±1,5% | 13 (11-18) 100,0% |
4 | 1 (1-2) 4,1±0,3% | 3 (2-5) 62,0±2,7% | 5 (4-5) 33,9±2,4% | 9 (7-13) 100,0% |
5 | 1 (1-3) 3,4±1,7% | 4 (4) 69,6±6,2% | 3 (4-3) 27,0±7,8% | 8 (8-9) 100,0% |
Примечание: в числителе количество компонентов, в скобках их размах колебаний, в знаменателе относительное содержание фракции в общем содержании белков мышц
изменения зафиксированы в белковом спектре мышц, имеющих высокую степень морфологических нарушений (Гераскин и др., 1990). По мере углубления мышечной патологии у осетра наблюдается нарастание белкового дефицита, снижение в крови холестерина, с тенденцией понижения у рыб старше 11 лет кортизола, при резко выраженной гиперкальцемии (Кузьмина и др., 1992; Наточин и др., 1995). У рыб с высокой степенью патологии мышц, выявлено также уменьшение (p<0,05) активности ЦО в печени более чем вдвое и увеличение в мышечной ткани в 2,6 раза, при почти неизменном уровне активности ЛДГ и СДГ. Активность ЛДГ в крови (67±11 мкмоль/мин·л) у таких рыб была в два раза выше, чем у особей с нормальными мышцами (35±5 мкмоль/мин·л). На уровне тенденции (на 20%) отмечено снижение активности АОА в печени, у рыб с нарушениями морфологической структуры мышц, что является результатом высокой интенсивности ПОЛ.
У севрюги чёткой связи между биохимическими показателями рыб и глубиной морфологических изменений в мышцах не прослеживалось, за исключением активности ЦО и АОА в печени. Причины таких различий кроятся в видовой специфике развития по времени и степени выраженности реакций организма севрюги в ответ на действие возмущающих факторов среды.
У белуги просматривалось некоторое углубление функциональных сдвигов у особей с дегенеративными изменениями мышечной ткани, как на уровне тенденций, так и подтверждаемые статистически.
Такое развитие реакции организма осетровых рыб на токсическое воздействие окружающей среды можно оценить как острый токсикоз - результат существенного повышения содержания в воде Северного Каспия нефтепродуктов, пестицидов и фенолов (Попова и др., 1992; Мумжу и др., 1989; Ежегодник … за 1990 г.). При его снижении в 1990 г., наблюдается уменьшение уровня функциональных и гистоморфологических изменений.
4. Влияние различных уровней загрязнения Каспийского моря на физиологические системы осетровых рыб
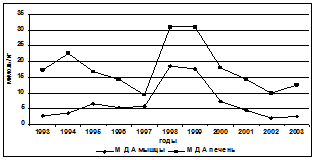
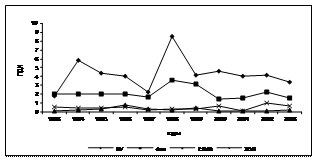
Введение в комплекс показателей, характеризующих физиологическое состояние осетровых, изучение процессов ПОЛ, выявило их значительную изменчивость, которая проявляла определённую зависимость от содержания в морской воде нефтепродуктов, что видно на примере русского осетра (рис. 2).
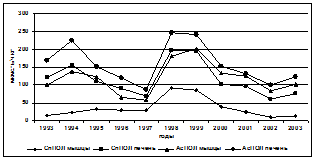
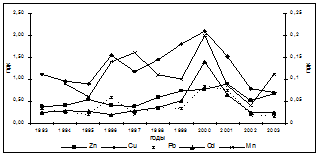
ХОП мкг/л Х 10 Zn, Cu, Pb – левая; Сd, Mn - правая шкала
Рис. 2. Межгодовая динамика интенсивности АсПОЛ и СпПОЛ, содержания МДА в печени и мышцах русского осетра и загрязнения моря (по данным лаборатории водных проблем и токсикологии КаспНИРХ) органическими поллютантами и некоторыми металлами
Другая закономерность, связана с сезонными колебаниями содержания в воде нефтяных углеводородов - преимущественным снижением их содержания от летнего максимума к осени (Глумов, 2001), которая отражалась и на сезонной динамике ПОЛ (табл. 4). Интенсификация ПОЛ в печени и мышцах летом зафиксирована у всех исследованных видов осетровых. Она обусловлена выносом токсических веществ паводковыми водами в море и увеличением температуры воды. Считается (Глумов, 2001), что основной объём (около 90%) загрязняющих веществ в Северный Каспий поступает с речным стоком, при сохранении примерного соотношения загрязняющих веществ.
Таблица 4
Параметры перекисного окисления липидов (М±m) мышц и печени русского осетра в разные сезоны года за исследуемый период ( гг.)
Сезоны года | СпПОЛ мкмоль/ч кг | АсПОЛ мкмоль/ч кг | МДА мкмоль/кг | |||
мышцы | печень | мышцы | печень | мышцы | печень | |
Весна | 28,2±4,0 | 75,4±6,2 | 113,6±7,6 | 108,2±8,8 | 6,2±1,0 | 10,6±0,8 |
Лето | 42,6±3,4 | 136,8±9,2 | 133,0±5,4 | 184,2±11,4 | 8,8±0,8 | 21,4±1,6 |
Осень | 24,2±2,0 | 95,8±7,2 | 111,2±4,4 | 142,0±10,2 | 4,6±0,4 | 16,0±1,2 |
Зависимость параметров ПОЛ русского осетра от уровня общего загрязнения вод Северного Каспия демонстрирует и регрессионный анализ. Коэффициент множественной корреляции в этом случае очень высок: от 0,84 до 1,00. Однако достоверными являются лишь регрессионные линейные уравнения зависимости скорости СпПОЛ (У1) в мышечной ткани и накопление в ней МДА (У2) от концентраций в воде исследованных токсикантов: У1 = - 0,20Х1 + 1,37Х2 + 0,416Х3 - 0,35Х4 + 0,44Х5 – 0,88Х6 + 0,21Х7 + 1,05Х8 + 0,249Х9 – 31,59; У2 = - 0,29Х1 + 1,49Х2 + 044Х3 – 0,31Х4 + 0,088Х5 – 0,98Х6 + 0,26Х7 + 1,07Х8 + 0,28Х9 -7,64 соответственно, где Х1 – нефтеуглеводороды; Х2 – фенолы; Х3 – СПАВ; Х4 – ХОП; Х5 – Zn; Х6 – Cu; Х7 – Pb; Х8 – Cd; Х9 – Mn. В этом случае коэффициент множественной корреляции достигает 1,00, показывая прямую линейную зависимость этих параметров ПОЛ от уровня общего загрязнения водной среды. Более детальный анализ выявил достоверную корреляционную связь ПОЛ с содержанием в воде нефтеуглеводородов, фенолов, а также меди и кадмия при высоких коэффициентах корреляции: от 0,61 до 0,84. В случае множественной регрессии, коэффициент корреляции уровня ПОЛ с этим поллютантам достигает 0,84. Из этого следует, что основу негативного влияния загрязнений на русского осетра составляют нефтеуглеводороды, фенолы и кадмий, а также, в некоторой степени, медь.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
