
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Анализ в этом отношении параметров ПОЛ двух других видов осетровых дал результаты, свидетельствующие о видовых особенностях реакции этих рыб на загрязнение среды обитания. Однако и в этом случае во всех вариантах регрессионного анализа, достоверно отражающих влияние компонентного состава загрязнения, на первом месте присутствуют нефтеуглеводороды и лишь как дополнение к ним тяжелые металлы и один или два вещества органического происхождения. То есть уровень ПОЛ у осетровых рыб обуславливают не отдельные поллютанты, а их комплекс, индуцируя состояние окислительного стресса. Во всех вариантах множественной регрессии ведущим элементом модели являются нефтеуглеводороды, затем фенолы, а потом тяжёлые металлы. Видовые особенности выражаются в проявлениях связи между параметрами ПОЛ и отдельными видами токсикантов или их сочетанием. Среди тяжёлых металлов выделяется кадмий, который, видимо, в сочетании с нефтеуглеводородами и фенолами увеличивает токсичность последних.
Реагирует на изменения загрязнения морских вод и энергетическая система осетровых. Её ответ проявляется изменениями активностей ключевых ферментов аэробного и анаэробного пути получения энергии в печени и мышцах - цитохромоксидазы (ЦО) и лактатдегидрогеназы (ЛДГ). Из-за того, что активность этих ферментов определялась летом, то и уровень загрязнения в сравнительном анализе использовался за это время года. А он, как показано выше, зависит от объёма паводка. При больших паводках активность ЦО в печени снижалась, низких - увеличивалась (рис. 3). Другим доказательством
![]()
![]()
![]()
![]()
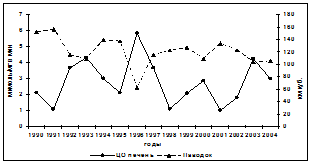
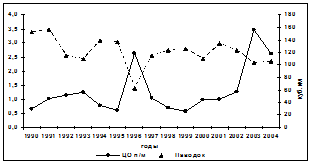
Рис. 3. Межгодовая динамика активности ЦО в печени (а) и соотношение (б) её активностей в печени и мышцах у русского осетра в летний период при различных объёмах паводка
влияния поллютантов на активность ЦО в печени является её уменьшение у особей осетра, выловленных в районах прямого действия волжского стока, более чем вдвое (р<0,05), в сравнении с рыбами, из других участков моря. Кроме того, изменяется также соотношение активностей ЦО в печени и мышцах (рис. 3). Если оно меньше единицы, то можно говорить о нарушении в распределении функциональных приоритетов ферментных систем, регулирующих аэробные и анаэробные процессы в энергетическом обмене печени и белых мышцах. Печени свойственна высокая активность ферментов аэробного обмена, а белым мышцам - анаэробного гликолиза (Щепкин, Эмеретли, 1985; Эмеретли, 1985; Иванов, 2003). При этом прослеживается видовая специфичность ответной реакции этого фермента в печени и мышцах на уровень загрязнений, вносимых паводковыми водами в Северный Каспий. У русского осетра она более замедлена, в сравнении с севрюгой и белугой.
Переходя к анализу динамик активности ЛДГ в печени и мышцах у осетровых рыб, необходимо отметить определённую их противофазность по отношению к динамике величины паводка (рис. 4) с проявлением видовой специфичности. У русского осетра такая закономерность проявлялась в большей степени в активности ЛДГ печени, но лишь до 2000 г., после которого характер динамики изменился. Однако, несмотря на видовые особенности, у
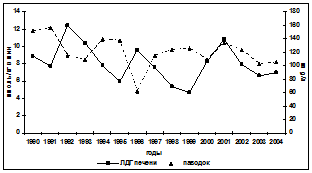
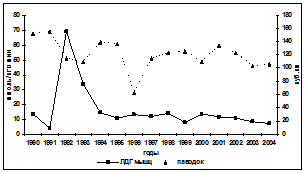
Рис. 4. Межгодовая динамика активности ЛДГ печени и мышц русского осетра
в летний период при различных объёмах паводка
всех исследованных видов осетровых выявляются общие закономерности. Успешное функционирование энергетической системы, зависит также и от работы дыхательной системы. Органом извлечения кислорода из водной среды у взрослых рыб являются жабры. Они чутко реагирует на присутствие в воде токсикантов. Гистологические исследования состояния жаберного эпителия у осетровых рыб, выловленных в море, выявили наличие различных форм нарушений, затрагивающих все отделы жаберного эпителия, независимо от пола, стадии зрелости и видовой принадлежности. При существенном увеличении как в 1996 г., содержания в морских водах меди, свинца и марганца, а также углеводородов, резко, до 51% выросла встречаемость рыб с тяжёлыми изменениями в жабрах при примерно 30% встречаемости в другие годы. Сильно повреждают жабры ионы цинка, который обладает синергическим действием в смеси с медью (Метелёв и др., 1971). Увеличение концентраций цинка и меди к 1999 г. также сказалось на состоянии жаберного эпителия - у 40% рыб изменения носили существенный характер. Кроме того, респираторный эпителий большинства рыб был разрыхлён, отмечалось его набухание и гиперплазия. В то время как до 1998 г., осетровым в море было характерно лишь разрастание эпителия филамента.
Таким образом, ведущим фактором в повреждении жабр выступает медь, токсичность которой может усиливаться цинком (синергический эффект), а так же совместным действием других металлов.
Другой компонент дыхательной системы рыб - гемоглобин, реагирует на загрязнение среды увеличением или снижением своего содержания в крови. У русского осетра оно синхронно следует за уровнем основных загрязнителей морских вод. У севрюги - в основном проявляется обратная зависимость. Такие различия связаны с разной стратегией адаптации организма осетра и севрюги и их этапностью к действию поллютантов. У осетра - повышением потребностей в окислительных процессах, у севрюги их торможением, наступающим в поздних фазах токсикоза. У белуги, картина реагирования гемоглобиновой системы схожа с таковой у севрюги, но с заметной инерционностью процессов.
Отклик регуляторно-гомеостатической функции на воздействие поллютантов рассматривался в свете изменений метаболизма, как регуляторно-компенсационных реакций. На воздействие чрезвычайных раздражителей, одним из первых реагирует углеводный обмен, который может быстро удовлетворить повышенные на этот момент потребности печени в энергии, что предусматривает расход гликогена. В иных случаях, когда гликоген не расходуется, как в и гг., организм рыб переходит на другую стратегию адаптации к действующим чрезвычайным факторам среды, используя в качестве энергетического субстрата другие вещества.
В качестве активного участника адаптационных процессов рассматривался также холестерин - основа стероидных гормонов, являющихся регуляторами обмена веществ в клетках. Содержание в крови холестерина реагирует на межгодовые изменения уровня поллютантов разнонаправленными по отношению к ним колебаниями, что наиболее выражено у русского осетра и севрюги. У белуги увеличение загрязнения каспийских вод не влекло за собой существенного снижения в крови холестерина, а сама динамика в гораздо большей степени, чем у осетра и севрюги, имела синхронность в отношении колебаний уровня загрязнения.
Бета-липопротеиды, представляющие собой транспортную форму липидов, переносят преимущественно холестерин и фосфолипиды. Севрюга, как наиболее реактивный вид, реагирует на повышенную загрязнённость вначале постепенным повышением содержания β-липопротеидов в крови, а затем резким снижением с поддержанием этого небольшого уровня в последующие годы. Те же процессы имеют место и у осетра, но без срывов на более низкий уровень. Белуга занимает промежуточное положение с чётко обозначенной реакцией изменения β-липопротеидов на динамику загрязнённости морских вод нефтеуглеводородами и фенолами.
Реакция белкового обмена осетровых на изменения уровня загрязнений в водной среде рассматривалась исходя из трёх показателей: содержания общего белка в крови, водорастворимого белка мышц и коэффициента, характеризующего уровень повреждения мышечных волокон, рассчитываемого как соотношение активностей ЛДГ в сыворотке крови и мышцах. Она представляет собой достаточно сложные взаимоотношения между деградацией мышечных волокон, белками мышц и крови и метаболической активностью печени. Прямой связи между межгодовыми изменениями уровня загрязнённости морских вод и содержанием в крови белков у русского осетра не прослеживается (рис. 5). Однако существенное повышение суммарного НУ+фенолы загрязнения как 1992 и 1998 гг. (10-12 ПДК) влечёт за собой снижение содержания белков в крови, как и длительный по времени (1994-
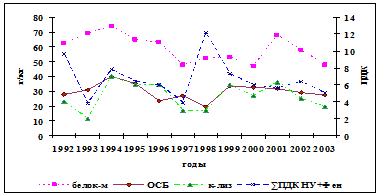
Рис. 5. Межгодовая динамика содержания в крови русского осетра общего сывороточного белка (ОСБ), водорастворимого белка мышц и коэффициента лизиса мышечных волокон
1996 гг.) повышенный (6-8 ПДК) их уровень. При этом просматривается сопряжённость между уровнем водорастворимых белков мышц и содержанием в крови белков: увеличение первых сопровождается повышением вторых, при росте коэффициент лизиса, свидетельствующего об усилении разрушения мышечных волокон. По-другому реагирует белковая система рыб во второй половине периода исследований. У севрюги реакция на увеличение уровня загрязнённости морской воды нефтеуглеводородами и фенолами проявляется уменьшением содержания белка в крови, при не чётко выраженной реакции водорастворимых белков и коэффициента лизиса мышечных волокон. У белуги просматривается тенденция повышения содержания белков в крови при увеличении загрязнённости морских вод этими поллютантами и сопряжённости межгодовых изменений коэффициента лизиса мышечных волокон и водорастворимого белка в пределах всего исследованного периода. Такая неоднозначность отклика белкового обмена на изменения воздействий на организм основных загрязняющих веществ диктуется различиями стратегий адаптационных возможностей исследуемых видов рыб и связана с особенностями этих воздействий (длительность, интенсивность) как в предшествующие годы, так и в анализируемый период.
Из систем организма рыб, осуществляющих защитные функции, исследовались две: антиоксидантная и иммунная. У всех исследованных видов осетровых рыб АОА в общих чертах соответствовала межгодовым колебаниям интенсивности ПОЛ в мышцах и печени и динамике уровня загрязнения морских вод. Однако в ряде случаев имело место несовпадение подъёма АОА и увеличения ПОЛ в этих тканях. В целом отмечается некоторое угнетение АОА при интенсивном и длительном воздействии на организм рыб поллютантов.
Другая система, выполняющая защитные функции, представлена иммунитетом. Исследование ЕИФ, характеризующего состояние иммунного статуса выявило у осетровых рыб наличие вторичного иммунодефицита: наибольший - у севрюги, наименьший - у белуги. Русский осётр занимал в этом отношении промежуточное положение. Несмотря на некоторые различия, у всех видов он оценивался как умеренный.
Таким образом, функционирование рассмотренных физиологических систем трансформируется в зависимости от силы и длительности воздействия и участия их в адаптационно-компенсационных процессах.
5. Формирование реакций организма осетровых рыб на загрязнение их среды обитания
В природной среде трудно зафиксировать не только начальные стадии интоксикационного процесса, но и его развитие. Однако, наряду с тщательным анализом данных, применение математических методов дает такую возможность. Детальный анализ исследованных показателей у осетровых рыб позволил выявить четыре периода, имеющих свои особенности отклика организма на загрязнение водной среды (табл. 5). Хотя, в целом, они
Таблица 5
Периоды в исследовании осетровых рыб, различающиеся ответными реакциями на воздействие загрязнений вод Каспийского моря
ПЕРИОДЫ | |||
I 1 половина 80-х | II 2 половина 80-х | III
| IV
|
НАРО - стойкая активация Прямое воздействие пестицидов на гонады | Стресс-реакция. Фазы: тревоги, резистентности (сопротивляемости) истощения (метаболической депрессии). Прямое воздействие пестицидов на гонады | Хроническая фаза: ухудшение, улучшение физиологического состояния | Фаза неустойчивого функционального равновесия |
Мобилизация энергоресурсов – нейтрализация и вывод из организма ксенобиотиков. Замедление темпов формирования гемоглобиновой системы, развития половых желёз. Эстрогенный эффект пестицидов - гормональные изменения. Нарушения: генеративного метаболизма; морфофункциональные гонад и ооцитов | Углубление изменений в метаболических процессах вовлечение в адаптивный процесс других физиологических систем. Заканчиваются море: гипометаболическое состояние. река – истощение и гибель Результирующая: наибольшее разнообразие и глубина нарушений в морфофункциональном состоянии тканей мышц, печени и гонад (в т. ч. эстрогенного эффекта пестицидов) | Снижение уровня патологичности мышц и гонад, стабилизация - в печени, некоторое ослабление защитно-приспособительных функций. Слабая гипоксия из-за снижения активности ЦО. Компенсируется активизацией ЛДГ и ЦО в мышцах, ЛДГ в печени частичное восстановление некоторых физиологических систем | Постепенное восстановление антиоксидантной системы, увеличение аэробной составляющей в энергетике печени, умеренный вторичный иммунодефицит. Использование мышечного белка для энергетических нужд, повышение антиоксидантной активности, глюконеогенез с увеличением гликогена печени. |
укладывались в концепцию общих неспецифических адаптационных реакций организма (НАРО) животных (Гаркави и др., 1990). Кроме того, с помощью кластерного анализа у русского осетра были выявлены 12 групп рыб, отражающих последовательность реакции организма на разные уровни и длительность воздействия поллютантов, среди которых особи, не испытывающие воздействие загрязнения. У них низкие величины параметров ПОЛ, на фоне высокой АОА, сдерживающий этот процесс в рамках физиологической целесообразности. Активности ЦО и ЛДГ сбалансированы и соответствовали особенностям функционирования энергетической системы печени и мышц, при достаточно высокой их активности. Ответ на невысокий уровень раздражающего фактора соответствовал различным фазам реакции спокойной (первичной) активации по классификации НАРО: рост антиоксидантной активности в печени и мышцах, усиление использования гликогена печени на энергетические нужды, при главенствующей роли аэробного обмена. Такие изменения у рыб можно расценивать как небольшая стимуляция обменных процессов, переходящая затем, в более выраженные изменения. Более высокий их уровень вызывал по классификации НАРО - реакции повышенной активации, проявившейся мобилизацией всех ресурсов организма (повышение Хл, Hb, ЦО, ЛДГ, снижение Гл печени, ОБ крови и ВРБ мышц). На увеличение длительности такой силы воздействия организм реагирует небольшим повышением АОА, большим снижением энергетического обмена в печени, в сравнении с мышцами, с уменьшением поступления кислорода и интенсивном использовании гликогена печени. Ответ физиологических систем осетра на дальнейший рост загрязнения моря проявляется как стресс-реакция, которая развивается в том же направлении, как и при меньших уровнях загрязнения. Стратегия организма в этот момент направлена на получение энергии любым путём, в короткие сроки из всех возможных источников: углеводов, белков, липидов и представляет собой реакцию тревоги, переходящая в стадию резистентности, а затем, в классическом представлении, истощения. Однако в природных условиях организм осетровых переключался на минимизацию метаболизма, о чём свидетельствует обеспечение энергией в основном анаэробным путём. При этом отмечается восстановление уровня гликогена в печени и высокий уровень ВРБ в мышцах, что в совокупности с тенденцией к нормализации показателей крови, дает основание говорить не об истощении, а о переходе в гипометаболическое состояние. У рыб, испытывающих сильный и сверхсильный стресс, очень высокие показатели ПОЛ, при низкой АОА, с мобилизацией срочного получения из всех источников и разными путями энергии, которая в большей степени генерировалась в мышцах, а не в печени. Описанные выше основные адаптивные реакции организма осетровых, нашли свое отражение в схеме (рис. 6), которая представляет предполагаемый
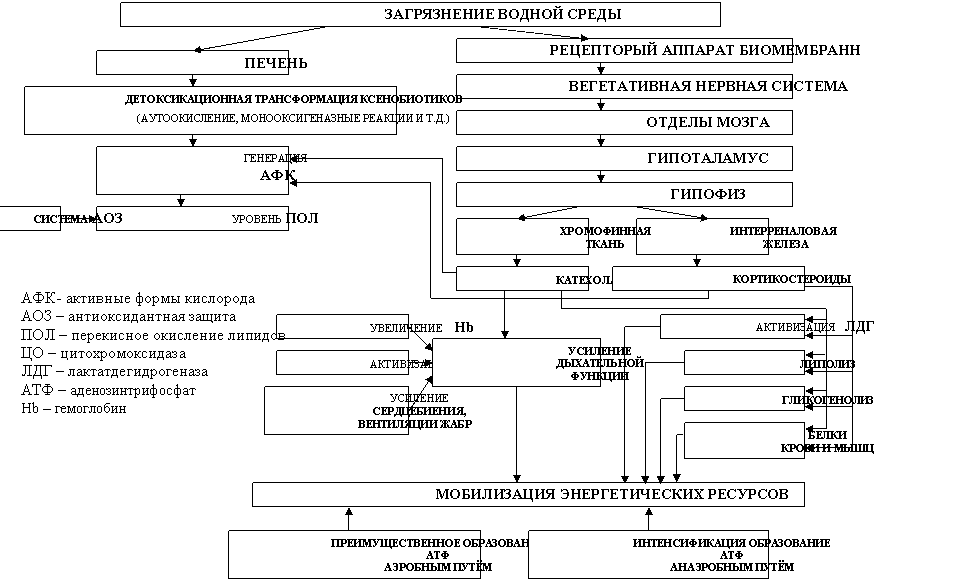

Рис. 6. Предполагаемый механизм первой фазы НАРО на загрязнение среды обитания осетровых рыб
механизм первой фазы НАРО на загрязнение водной среды - процесс срочного получения энергии, при балансе ПОЛ и АОА.
Дальнейшее развитие реакций организма на загрязнение воды сопровождается ростом интенсивности ПОЛ при снижении АОА и перестройкой энергетического обмена с усилением анаэробного пути получения энергии, вследствие снижения активности ЦО. Считается (Кожевников, 1985; Балаболкин, 2000), что ингибиторами ЦО являются активные радикалы и в частности оксид азота (Xie et al., 1996; Balakirev et al., 1997), который способен конкурентно связываться с этим ферментом (Bates et al., 1996). Ингибирование цитохромоксидазы, отмечаемое у осетровых рыб, с увеличением интенсивности ПОЛ и снижении АОА, по всей видимости, имеет тот же механизм. Как видно из рисунка 7 активность цитохромоксидазы с увеличением интенсивности перекисных процессов возрастает лишь при небольших уровнях ПОЛ, а затем начинает снижаться до минимальных значений.
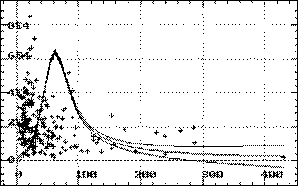
Рис. 7. Регрессионная модель активности цитохромоксидазы от уровня АсПОЛ
Такая динамика отношений параметров ПОЛ и активности ЦО предполагает фазность функционирования этого фермента: до нарушения баланса между образованием и инактивацией продуктов ПОЛ антиоксидантами
и после. Как только нарушается баланс между образованием и нейтрализацией продуктов ПОЛ антиоксидантами, стрессорная реакция переходит в новую фазу, где стратегия адаптации к сильному, либо длительно действующему или часто повторяющему фактору существенно изменяется (рис. 8). Она направлена на снижение образования свободных радикалов и подавления их активности, при понижении метаболизма до предельно низкого уровня. Реализуется за счёт получения энергии преимущественно бескислородным путём (снижение доступа кислорода к клеткам), а также восстановления антиоксидантной активности. Одним из основных путей возможной компенсации недостатка АТФ в условиях эндогенной гипоксии, вызванной окислительным стрессом, является эволюционно закрепленное у осетровых рыб получение энергии из мышечных белков во время нерестовой миграции (при переходе на эндогенное
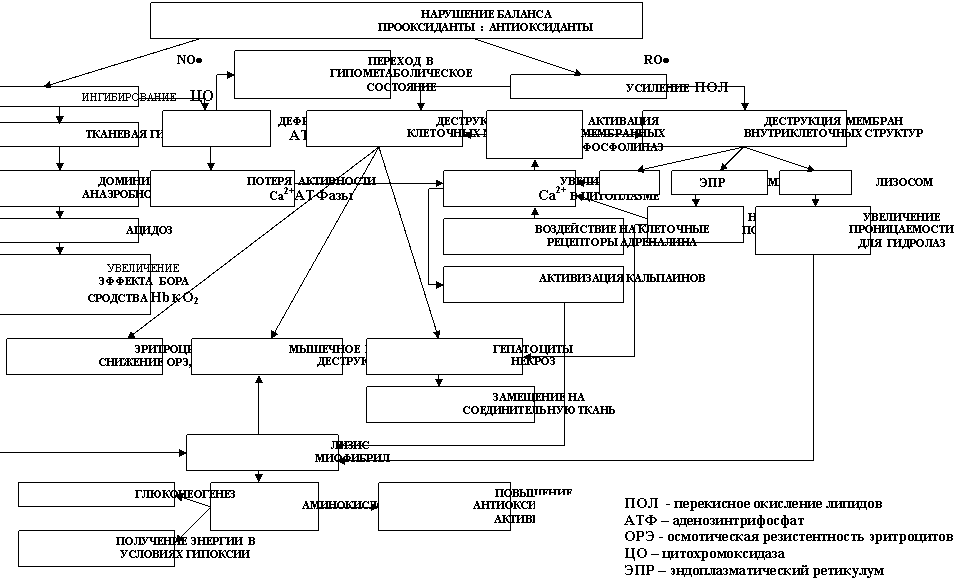

Рис. 8. Предполагаемый механизм второй фазы НАРО на загрязнение среды обитания осетровых рыб
питание). В процессе анаэробного катаболизма белков (Шульман и др., 1993) освобождается достаточное количество энергии для ресинтеза АТФ, что позволяет осетровым выживать при гипометаболическом состоянии. В деградации миофибрилярных белков до аминокислот, наряду с кальпаинами участвуют и катепсины (Немова, 1992), при возможном участии и других протеолетических ферментов. Тем самым создаются условия для усиления антиокислительной защиты, за счет аминокислот и образования глутатиона, предотвращая дальнейшее развитие окислительного стресса под воздействием ксенобиотиков, что подтверждается экспериментами на теплокровных животных (Штаркман, 2008; Кулинский, Колесниченко, 1990).
Таким образом, можно говорить, что деструктивные изменения или иначе «расслоение» мышц у осетровых рыб обусловлены тканевой или точнее гепатотоксической гипоксией (Гераскин, 1989). Это подтверждается как прямыми опытами с токсикантами на молоди русского осетра и белуги (Алтуфьев и др., 1989; Алтуфьев, 1994), так и более поздними публикациями (Евгеньева, Семёнова, 2000; Ланге, Кловач, 2002; Зайцева, 2002; Алтуфьев, Гераскин, 2003). Одним из признаков токсического поражения печени является жировая инфильтрация переходящая в жировую дистрофию (Лейтес, Лаптева, 1967; Логинов, Аруин, 1985), что отмечалось у каспийских осетровых в гг.
ЗАКЛЮЧЕНИЕ
Реакция осетровых на загрязнение токсичными для рыб веществами, зависит от силы и длительности их действия и проявляется в последовательном вовлечении в адаптационный процесс необходимых для этого функциональных систем. В условиях естественной среды, развитие токсического процесса протекает одновременно с мобилизацией адаптивных реакций к действующему фактору, которые обладают более выраженным эффектом.
В основе функциональных преобразований, происходящих в организме рыб под действием загрязнений и реализации детоксикационной функции, лежат два базовых процесса - баланс прооксидантов и антиоксидантов и удовлетворение всеми возможными путями повышенных потребностей на данный момент в энергии. При низких и умеренных уровнях загрязнений среды и небольшой длительности их действия, антиоксидантная система способна гасить активность свободных радикалов, появляющихся при действии ксенобиотиков, а уровень отклика соответствует по классификации НАРО этапам спокойной и повышенной активации. На этих этапах отмечается нарастание активности метаболизма, направленного на получение быстрого и максимального количества энергии. Главную роль в обеспечении организма энергией в это время играет аэробное окисление, несмотря на увеличение анаэробного. Невысокий уровень загрязнения среды, но при большой длительности их действия, приводит к уменьшению АОА и постепенному снижению роли аэробного обмена и повышении анаэробного в получении АТФ. В условиях умеренной и высокой силы, роль печени в обеспечении энергией организма снижается, а мышц - повышается. При этом проявляются видовые особенности, связанные с различной реактивностью рыб. При значительном уровне или длительности воздействия на рыб токсикантов реакция организма осетровых развивается как стресс-реакция, со всеми свойственными для неё фазами, и заканчивается гипометаболическим состоянием. В этом случае отмечается истощение АОА и нарушение баланса прооксиданты : антиокисданты в пользу первых, уменьшение аэробной составляющей в получении энергии организмом, из-за снижения активности цитохромоксидазы, ингибируемой, по всей видимости, радикалами нитрооксида. Изменяется и стратегия адаптивных реакций, которая направлена на снижение образования свободных радикалов и подавления их активности, при понижении метаболической функции до предельно низкого уровня. Она реализуется за счёт получения энергии преимущественно бескислородным путём (снижение доступа кислорода к клеткам), при анаэробном окислении продуктов деградации мышечных белков, а также увеличения антиоксидантной активности, которой обладают некоторые аминокислоты и трипептид глутатион. Механизм повреждения клеток универсален и сводится при интенсификации ПОЛ к повреждениям липидного биослоя мембран органоидов и клеток, сопровождающихся нарушением митохондриального окисления и каскадом метаболических нарушений в форме тканевой гипоксии, ацидоза, увеличения Са2+, активизацией гидролитических ферментов, деградацией метаболизма и гибелью клетки. На макроскопическом уровне эти процессы проявляются некрозом ткани и развитием фиброза и заменой на жировую в печени и деструкцией волокон мышц.
В целом изменения, выявляемые в организме осетровых рыб, при загрязнении окружающей среды соответствуют признакам гепатотоксической гипоксии, одной из форм тканевой гипоксии, представляющей собой токсическое поражение печени, при снижении поступления в гепатоциты кислорода из-за ингибирования активности цитохромоксидазы. Одним из признаков токсического поражения печения является жировая инфильтрация, переходящая в жировую дистрофию, отмечаемая у осетровых рыб.
Не менее существенными являются последствия ухудшения физиологического состояния осетровых, выражающиеся в снижении темпа роста и воспроизводительного потенциала, связанного с удлинением времени вступления в репродукционную стадию и межнерестовых периодов, а также патологических изменений, сопровождающие гонадо - и гаметогенез осетровых, что подтверждается данными ихтиологических исследований (Журавлёва, 2005) констатирующих снижение темпа роста и индивидуальной плодовитости. Особую тревогу вызывает уменьшение плодовитости связанное с нарушениями в половых железах и, особенно, замена генеративной ткани на жировую или соединительнотканную. Такой же процесс выявлен и у самцов. Снижению воспроизводительного потенциала способствуют и другие формы патологий ооцитов. Они проявляются: не оплодотворением икры, гибели её в процессе эмбриогенеза, а также повышенной элиминации на ранних этапах развития и снижения резистентности молоди к экстремальным факторам внешней среды (Лагунова, 1997; Лепилина, Шевелева, 1997; Шагаева и др., 1989). На данном этапе последствия изменений в физиологическом состоянии осетровых рыб пока лишь ограничивают биопродуктивность этого семейства. Однако дальнейшее развитие этого процесса может привести к критическому состоянию, в понимании и (2005), обитающих в Каспийском море популяций осетровых рыб при превышении порога устойчивости рыб к повреждающим факторам внешней среды.
ВЫВОДЫ
1. Начальные признаки реакции организма осетровых рыб на загрязнение водной среды в большей мере проявляются у самок в наиболее чувствительный для них преднерестовый период нарушением вителлогенеза, истощением плохо подготовленных к нересту рыб и морфофункциональными отклонениями в развитии ооцитов. В морской период жизни - на уровне более тонких функциональных изменений, связанных с работой энергетической системы. У молодых особей - задержкой в развитии и становлении отдельных функций.
2. Развитие реакции организма осетровых на нарастание антропогенного воздействия характеризуется углублением изменений в физиолого-биохимических показателях и гистоморфологических структурах внутренних органов как в периоды физиологически спокойного состояния (II СЗГ) разновозрастных особей нагуливающихся в море, так и репродуктивной активности (III-IV СЗГ) связанной с созреванием гонад. Оно выразилось существенными изменениями в энергетическом и водно-солевом обменах, обмене веществ, нарушениями в морфофункциональном состоянии печени, мышечной ткани и гонад и соответствовали в своей основе картине развития тканевой или точнее гепатотоксической гипоксии. Интегральным выражением этих изменений стал феномен «расслоения» мышечной ткани.
3. Развитие токсического процесса у осетровых в природных условиях сопровождается рядом неспецифических адаптивных реакций к загрязнению среды обитания при их высокой выраженности, вследствие чего действие загрязняющих веществ в различном сочетании вызывают однотипные реакции организма. Установлено прямое влияние (r=1) уровня общего загрязнения вод Северного Каспия, на интенсивность перекисного окисления липидов у осетровых рыб, описываемое уравнениями множественной регрессии.
4. Обнаруженные различия в реакциях исследованных физиологических систем, на воздействие токсических веществ в рассматриваемые годы, дают основание выделить четыре периода, имеющих свои особенности отклика на эти воздействия. Первому характерны признаки стойкой (повышенной) активации согласно общей теории адаптивных реакций. Второму - острого отравления и развития стресс-реакции во всех трёх её стадиях. Третий этап - переход к хронической фазе. Четвертому - характерен хронический токсикоз с переходом на иную, чем в третьем, стратегию адаптационно-компенцационных реакций.
5. Выявленные функциональные изменения у осетровых рыб представляют собой комплекс неспецифичных реакций адаптаций, степень вовлечённости в которые физиологических систем зависит от силы и длительности воздействия на рыб токсичных для них веществ и представлены по классификации НАРО от стадий спокойной (первичной) и стойкой (повышенной) активации до стресс-реакции, заканчивающейся чаще всего метаболической депрессией.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
