


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
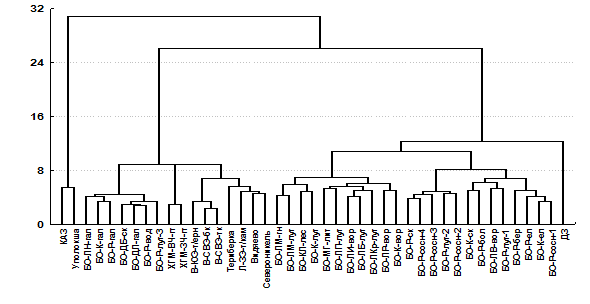
Рис. 11. Дендрограмма сходства природных и техногенных биотопов по видовому составу орибатид (Ward`s method, Euclidean distances). БО – биотопы Беломорских островов.
В импактной зоне медно-никелевого предприятия, функционирующего в таежной зоне области, комплекс орибатид с крайне обедненной таксономической структурой был ближе к зональным и горным тундрам. Это свидетельствует о большем стрессирующем воздействии фактора техногенной трансформации почв на разнообразие орибатид по сравнению с экстремальными природно-климатическими условиями Кольского Севера.
6. ПАНЦИРНЫЕ КЛЕЩИ ТЕХНОГЕННЫХ СУБСТРАТОВ
(НА ПРИМЕРЕ ОТХОДОВ АПАТИТ-НЕФЕЛИНОВОЙ ПРОМЫШЛЕННОСТИ)
Обсуждаются результаты исследования пионерных фаунистических комплексов, которые сформировались в самозарастающих и рекультивированных нефелинсодержащих песках – отходах обогащения апатит-нефелиновых руд, складированных на хранилищах 0-40 лет назад и представляющих техногенный субстрат, отличный от зональных почв.
Первопоселенцами техногенного субстрата являются микробофаги-коллемболы, что объясняется наличием жизнеспособных бактериальных клеток в минеральных отходах обогащения апатит-нефелиновых руд (Евдокимова, Калмыкова, 2010). В отвалах 10-30 летнего срока складирования доля орибатид варьировала от 4% (100-200 экз./м2) в песках чистых и самозарастающих мхами до 20-35% (5.5 тыс. экз./м2) в ризосфере злаков, при посеве которых (для закрепления песков) вносили торф.
Формирование органогенного горизонта и растительного покрова в результате рекультивации и с увеличением срока складирования способствовало повышению микробиологической и ферментативной активности в песках (Евдокимова, Калмыкова, 2010), росту плотности панцирных клещей, их более равномерному пространственному распределению (особенно под мхами и лишайниками с более стабильным режимом увлажнения), смене жизненных форм, различиям в сроках и интенсивности репродукции. В песках, закрепленных > 40 лет назад, в составе микробиоты по биомассе преобладали микромицеты, а доминантами фауны стали панцирные клещи, связанные с грибами трофически. Численность клещей была сопоставима с уровнем зональных подзолов: 16-37 тыс. экз./м2 (рис. 12). Различия в численности орибатид в песках, закрепленных 10-30 лет назад и рекультивированных > 40 лет назад, составляли в разные месяцы вегетационного сезона от 9 до 110 раз (достоверно). Достоверного роста плотности коллембол с увеличением срока складирования песков не выявлено.
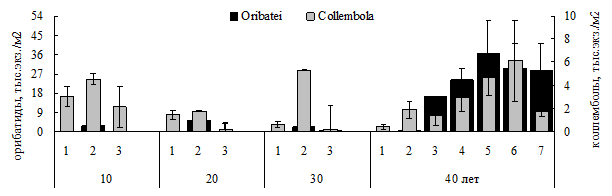
Рис. 12. Численность коллембол и орибатид в песках разного срока складирования. 1 – чистые пески, 2 – злаки, 3 – мхи, 4 – лишайники, 5 – вороника, 6 – брусника, 7 – клевер.
Установлена высокая корреляция плотности орибатид с ферментативной и микробиологической активностью в песках разного срока складирования (с биотическими факторами) и менее сильная – с их физико-химическими показателями (абиотическими факторами). Положительные корреляции с численностью микромицетов (r = 0.92), сапротрофных бактерий (0.77), активностью каталазы, фосфатазы, уреазы, инвертазы (r 0.83-0.92) и влажностью песков (0.50) отражали увеличение микробиологической и ферментативной активности и повышение влагоемкости под растительностью в «старых» отходах по сравнению с песками меньшего срока складирования. Отрицательные корреляции с температурой и pH водной суспензии объяснялись слабым прогревом песков под растительностью и снижением значений pH исходно щелочного субстрата до 6.7 под воздействием гумусовых кислот и продуктов жизнедеятельности биоты. Таким образом, в процессе первичного почвообразования на техногенном субстрате – отходах нефелинсодержащих пород панцирные клещи проявили себя как индикаторы микологической активности субстрата и накопления органического вещества.
7. ОБОБЩЕННАЯ ХАРАКТЕРИСТИКА
ФАУНЫ ПАНЦИРНЫХ КЛЕЩЕЙ МУРМАНСКОЙ ОБЛАСТИ
Обобщены данные по экологическому разнообразию (тип ареала, трофическая специализация, жизненные формы) для 259 видов панцирных клещей, выявленных на территории Мурманской области. Рассмотрено влияние фактора широтной зональности.
Показано преобладание видов с голарктическими (52% видов) и палеарктическими (38%) ареалами, как и на Европейском Севере России в целом (Мелехина, 2009). Малой мощности подстилки и укороченному профилю зональных почв соответствует высокая доля поверхностно-обитающих видов (50%), 30% приходится на обитателей почвенных скважин, 20% - на обитателей толщи подстилки.
Основу региональной фауны формирует эволюционно более молодая и прогрессивная группа высших панцирных клещей (Brachypylina), разнообразие низших орибатид (Macropylina) примерно в 2,5 раза ниже на уровне видов и надвидовых таксонов – родов и семейств (табл. 4).
Таблица 4. Соотношение высших (Brachypylina) и низших (Macropylina) орибатид
в таежных и тундровых почвах Мурманской области
Орибатиды | Область в целом | Северная тайга | Зона тундры | ||||||
Сем | Род | Вид | Сем | Род | Вид | Сем | Род | Вид | |
Всего, в т. ч.: | 53 | 113 | 258 | 47 | 102 | 216 | 38 | 78 | 142 |
Brachypylina | 39 | 83 | 182 | 33 | 74 | 154 | 29 | 60 | 101 |
Macropylina | 14 | 30 | 76 | 14 | 28 | 62 | 9 | 18 | 41 |
Соотношение, разы | 2,8 | 2,6 | 2,4 | 2,4 | 2,6 | 2,5 | 3,2 | 3,3 | 2,5 |
Треть семейств (34%) представлена единственным видом, 26% включают по 2-3 вида. Многовидовые таксоны Brachychthoniidae, Oppiidae, Suctobelbidae, Damaeidae, Ceratozetidae с числом видов от 15 до 18 составляют лишь 9%, объединяя при этом 30% видов. Клещи трех первых семейств – мелкоразмерные почвенные скважники, микрофитофагами (потребители грибов, бактерий, водорослей). Виды сем. Damaeidae и Ceratozetidae обитают на поверхности почвы и в толще подстилки. Видовая насыщенность семейств орибатид в исследованных районах приведена в табл. 5.
Таблица 5. Видовая насыщенность семейств орибатид в исследованных районах | |||||||||||
№ | Семейство / Район | 1 | 2 | 3 | 5 | 6 | 8 | 9 | 10 | 11 | |
Низшие орибатиды (Macropylina) | 1 | Paleacaridae (Grandjean 1932) | 2 | 1 | |||||||
2 | Brachychthoniidae (Thor 1934) | 10 | 5 | 14 | |||||||
3 | Hypochthoniidae (Berlese 1910) | 1 | 1 | 1 | 1 | ||||||
4 | Eniochthoniidae (Grandjean 1947) | 1 | 1 | ||||||||
5 | Parhypochthoniidae (Grandjean 1932) | 1 | |||||||||
6 | Eulohmanniidae (Grandjean 1931) | 1 | 1 | ||||||||
7 | Phthiracaridae (Perty 1841) | 1 | 3 | 3 | 1 | 3 | 1 | 3 | 7 | ||
8 | Euphthiracaridae (Jacot 1930) | 1 | 2 | 1 | 2 | 2 | 6 | ||||
9 | Malaconothridae (Berlese 1916) | 2 | 2 | ||||||||
10 | Trhypochthoniidae (Willmann 1931) | 2 | 1 | 1 | 1 | 1 | |||||
11 | Nothridae (Berlese 1896) | 1 | 3 | 1 | 1 | 3 | 6 | ||||
12 | Camisiidae (Oudemans 1900) | 2 | 7 | 2 | 6 | 2 | 2 | 8 | 10 | ||
13 | Nanhermanniidae (Sellnick 1928) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | |
14 | Hermanniidae (Sellnick 1928) | 1 | 1 | 3 | 1 | 3 | |||||
Высшие орибатиды (Brachypylina) | 15 | Hermanniellidae (Grandjean 1934) | 1 | ||||||||
16 | Licheremaeidae (Grandjean 1931) | 1 | |||||||||
17 | Damaeidae (Berlese 1896) | 1 | 9 | 3 | 2 | 2 | 3 | 11 | |||
18 | Cepheidae (Berlese 1896) | 1 | 1 | 1 | |||||||
19 | Eremaeidae (Sellnick 1928) | 1 | 1 | 1 | 1 | 2 | |||||
20 | Tenuialidae (Jacot 1929) | 1 | |||||||||
21 | Astegistidae (Balogh 1961) | 1 | 1 | 2 | 4 | ||||||
22 | Gustaviidae (Oudemans 1900) | 1 | |||||||||
23 | Liacaridae (Sellnick 1928) | 2 | 3 | 1 | 2 | 2 | 6 | 8 | |||
24 | Peloppiidae (Balogh 1943) | 2 | 1 | 1 | 1 | 3 | |||||
25 | Carabodidae (C. L. Koch 1837) | 3 | 3 | 5 | 1 | 4 | 6 | 3 | 3 | 9 | |
26 | Tectocepheidae (Grandjean 1954) | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 3 | ||
27 | Quadroppiidae (Balogh 1983) | 1 | 1 | ||||||||
28 | Caleremaeidae (Grandjean 1965) | 1 | |||||||||
29 | Oppiidae (Grandjean 1954) | 1 | 2 | 10 | 1 | 3 | 2 | 5 | 12 | 12 | |
30 | Suctobelbidae (Jacot 1938) | 1 | 1 | 6 | 1 | 9 | 7 | 1 | 8 | ||
31 | Autognetidae (Grandjean 1960) | 1 | 1 | 1 | 1 | 2 | 4 | 3 | |||
32 | Thyrisomidae (Grandjean 1954) | 2 | 3 | 3 | |||||||
33 | Hydrozetidae (Grandjean 1954) | 1 | |||||||||
34 | Limnozetidae (Grandjean 1954) | 3 | 2 | ||||||||
35 | Ameronothridae (Willmann 1931) | 4 | 6 | 6 | |||||||
36 | Cymbaeremaeidae (Sellnick 1928) | 1 | 1 | ||||||||
37 | Micreremidae (Grandjean 1954) | 2 | |||||||||
38 | Passalozetidae (Grandjean 1954) | 1 | |||||||||
39 | Scutoverticidae (Grandjean 1954) | 1 | 1 | 2 | 2 | ||||||
40 | Haplozetidae (Grandjean 1954) | 1 | |||||||||
41 | Phenopelopidae (Petrunkevitch 1955) | 1 | 5 | 1 | 1 | 1 | 1 | 4 | |||
42 | Achipteriidae (Thor 1929) | 1 | 1 | 1 | 1 | 2 | 2 | ||||
43 | Tegoribatidae (Grandjean 1954) | 1 | 1 | ||||||||
44 | Oribatellidae (Jacot 1925) | 1 | 3 | ||||||||
45 | Galumnidae (Jacot 1925) | 4 | 1 | 1 | 1 | 5 | 5 | ||||
46 | Ceratozetidae (Jacot 1925) | 2 | 2 | 10 | 5 | 1 | 8 | 11 | |||
47 | Chamobatidae (Thor 1938) | 1 | 1 | 4 | 1 | 1 | 1 | 1 | 2 | 2 | |
48 | Humerobatidae (Grandjean 1970) | 1 | |||||||||
49 | Mycobatidae (Grandjean 1954 | 1 | 1 | 1 | |||||||
50 | Euzetidae (Grandjean 1954) | 1 | |||||||||
51 | Parakalummidae (Grandjean 1936) | 2 | |||||||||
52 | Scheloribatidae (Grandjean 1953) | 1 | 2 | 5 | 1 | 3 | 1 | 3 | 5 | 5 | |
53 | Oribatulidae (Thor 1929) | 1 | 2 | 2 | 1 | 1 | 2 | 3 | |||
Видов в каждом районе | 14 | 27 | 123 | 12 | 42 | 38 | 41 | 113 | 178 | ||
Многовидовые семейства формируют основу фауны орибатид и в северотаежных, и в тундровых почвах области, однако для 9 из них в тундре число видов на 4-6 меньше, чем в тайге (рис. 13). Общими для Кольской тундры и тайги являются 48 видов (< 20% региональной фауны) из 23 сем. Степень фаунистического сходства выше на уровне семейств (Is = 38%), чем на видовом уровне (68%), что отражает большую специализацию видов к условиям среды по сравнению с таксонами высших рангов.
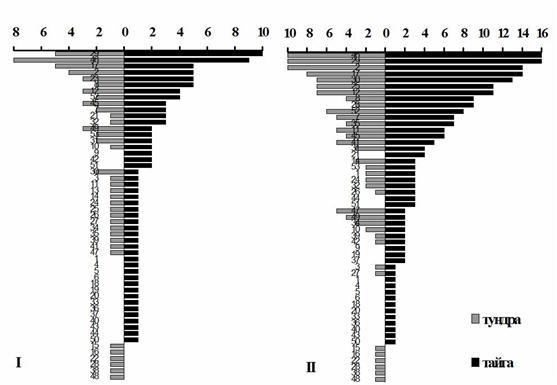
Рис. 13. Насыщенность семейств орибатид видами (I) и родами (II) в северотаежных и тундровых почвах Мурманской области. Обозначение семейств с 1 по 53 – как в табл. 5.
Сравнение списков орибатид северной тайги, тундры Мурманской области и граничащей с юга республики Карелия (северная и средняя тайга, 61-66° с. ш.) выявило тенденцию уменьшения числа одно - и многовидовых семейств с увеличением широты местности (табл. 6). В области отсутствуют 7 семейств, представленных в Карелии единственным видом, но есть 7 моновидовых таксонов, не указанных для республики.
Таблица 6. Соотношение семейств орибатид с разным числом видов
в почвах Мурманской области и республики Карелия
Семейства | Карелия, средняя тайга (Ласкова, 2001) | Мурманская область | ||||||
Подзона северной тайги | Зона тундры | Область в целом | ||||||
Сем. | Доля, % | Сем. | Доля, % | Сем. | Доля, % | Сем. | Доля, % | |
Одновидовые | 18 | 37 | 13 | 28 | 11 | 31 | 18 | 34 |
2-3 видовые | 14 | 29 | 17 | 36 | 9 | 25 | 14 | 26 |
≥ 4 видов | 17 | 35 | 17 | 36 | 15 | 42 | 21 | 40 |
Всего сем. | 49 | 100 | 47 | 100 | 36 | 100 | 53 | 100 |
Высокая доля одновидовых семейств орибатид в фауне северных регионов отражает миграционный характер ее формирования. Для Мурманской области распространение клещей возможно как минимум тремя путями: с морскими течениями (Нордкапская ветвь Гольфстрима), антропогенным привносом, путем форезии на мигрирующих птицах. Это подтверждается специфичностью видового состава и высокой долей локально встречающихся видов на морских побережьях. Из 37% локально встречающихся видов региональной фауны 24% (или 63 вида) найдены на побережье Баренцевого моря, 7% (17 видов) – на о-вах Белого моря, тогда как в горном районе – лишь 2% или 6 видов. Степень видового сходства выше для морских побережий (Is = 41%), чем для побережий и центральных районов (Is = 33-38%). Без учета локально встречающихся видов фауна орибатид региона насчитывает 162 вида.
«Прогнозное» разнообразие орибатид Мурманской области, рассчитанное по 11 исследованным районам, оценивается в 290 видов, что сопоставимо с разнообразием этих клещей для стран Северной Фенноскандии и превышает известное число видов для республик Карелия и Коми (Lundqvist, 1987; Schatz, 2004; Ласкова, 2001; Мелехина, 2004).
ВЫВОДЫ
1. Фауна естественных (равнинные и горные) и техногенно трансформированных северотаежных экосистем Мурманской области включает 216 видов панцирных клещей, принадлежащих к 102 родам, 47 сем. Список орибатид Кольской тундры расширен до 142 видов из 78 родов, 38 сем., общий региональный список – до 259 видов из 113 родов, 53 сем. В зоогеографическом отношении преобладают виды с голарктическими (52%) и палеарктическими (38%) ареалами; 50% видов представлены поверхностно обитающими формами; 70% видов орибатид являются панфитофагами.
2. Многовидовые таксоны Brachychthoniidae, Oppiidae, Suctobelbidae, Damaeidae, Ceratozetidae с числом видов от 15 до 18 объединяют треть видов региональной фауны. Высокая доля одновидовых семейств (34%) отражает миграционный характер формирования фауны орибатид Мурманской области, причем разными путями: форезией клещей на птицах, распространением с морскими течениями, антропогенным привносом.
3. Влияние широтной зональности на разнообразие панцирных клещей в пределах Мурманской области выражается в сокращении числа видов, родов и семейств, главным образом за счет таксонов низших орибатид (Macropylina), и в снижении доли двух-, и трехвидовых семейств при переходе от таежной зоны к тундровой.
4. В пределах таежного, субальпийского и альпийского высотных поясов Хибинского горного массива выявлено 49 видов орибатид из 37 родов, 23 сем. Семейство Haplozetidae и виды Protoribates capucinus, Heminothrus septentrionalis, Nanhermannia sellnicki обнаружены только в почвах Хибин. Установлено большее фаунистическое сходство горно-тундровых и горно-лесотундровых сообществ по сравнению с предгорными (горно-таежными). Предгорные леса на окраинах Хибинского массива, расположенного в северотаежной подзоне, ближе к зональным тундрам по сравнению с зональной тайгой.
5. В естественных экосистемах Кольского Севера с наиболее экстремальными природными условиями (зональные и горные тундры) и зонах сильного техногенного воздействия (медно-никелевый комбинат) формирование комплексов орибатид подчиняется общей закономерности: сокращение числа семейств и их обеднение до одного рода с единственным видом. При совпадении семейств, представленных в экстремальных природных и техногенно нарушенных экосистемах, сообщества орибатид имеют своеобразный набор видов.
6. Воздействие фтор-содержащих выбросов алюминиевого завода на почвенную фауну менее губительно по сравнению с выбросами медно-никелевого комбината. В зоне максимального загрязнения сохраняется высокое разнообразие орибатид, сопоставимое с зональной тайгой, и полидоминантная структура их сообщества. О негативном воздействии аэротехногенных выбросов на почву свидетельствует преобладание мелких эуэдафических видов из экологической группы почвенных скважников и супердоминирование эврибионтного партеногенетического вида O. nova.
7. Выделены категории панцирных клещей, устойчивых и чувствительных к промышленному загрязнению почв Мурманской области; среди них 13 представителей сем. Suctobelbidae и Carabodidae, преобладающих по числу видов в региональной фауне. Для индикаторных видов в загрязненных подстилках значимыми факторами являются численность и биомасса микроорганизмов, уровень кислотности, содержание углерода и соотношение C:N в подстилках.
8. Установлена сукцессия пионерных групп микроартропод, связанная со сменой бактериального и грибного компонентов микробиоты, в процессе первичного почвообразования на нефелинсодержащих отходах обогатительных фабрик. Увеличение срока складирования отходов сопровождается ростом численности орибатид, их более равномерным пространственным распределением, снижением доли мелкоразмерных обитателей почвенных скважин и возрастанием количества обитателей подстилки и поверхности почвы, что соответствуют перестройкам в сообществах панцирных клещей в ходе восстановительных сукцессий на нарушенных территориях.
Список публикаций по теме исследования
Публикации изданиях, рекомендованных ВАК
1. , , Пионерная фауна отвалов нефелин-содержащих пород // Почвоведение, 2009. № 8. С. 1-8.
2. , , Почвообитающие панцирные клещи (Acarina, Oribatei) таежной и тундровой зон Мурманской области // Зеленый пояс Фенноскандии / Труды Карельского научного центра РАН, 2011. № 1. Сер. Биогеография. Вып. 11. С. 54-67.
Публикации в тематических сборниках и трудах конференций
1. , А. Пионерная фауна нефелинсодержащих отвалов как показатель процессов первичного почвообразования // Экологические проблемы Северных регионов и пути их решения: Матер. Всерос. конф. Апатиты: Кольский НЦ РАН, 2008. С. 87-92.
2. , А. Сукцессия микроартропод в отвалах действующих и законсервированных хранилищ // Северные территории России: проблемы и перспективы развития: Матер. Всерос. конф. Архангельск: УрО РАН, 2008. С. 486-490.
3. А., Зенкова изученности почвообитающих панцирных клещей (Acarina, Oribatei) на территории Мурманской области // Актуальные проблемы сохранения биоразнообразия в экстремальных условиях северного климата: Матер. межд. науч. конф. Кировск: ПАБСИ РАН, 2008. С. 52-55.
4. А., Зенкова фауна нефелинсодержащих отвалов как показатель процессов первичного почвообразования // Почвенные сообщества: от структуры к функциям: Тез. XV Всерос. совещ. по почв. зоол. М.: ИПЭЭ, 2008. С. 284-287.
5. , А. Разнообразие панцирных клещей (Acariformes, Oribatei) в почвах Мурманской области // Сохранение биологического разнообразия наземных и морских экосистем в условиях высоких широт: Матер. межд. науч.-практ. конф. Мурманск: МПГУ, 2009. С. 96-101.
6. , А. Разнообразие панцирных клещей (Acariformes, Oribatei) в целинных и антропогенно нарушенных почвах Мурманской области // Продуктивность и устойчивость лесных почв: Матер. докладов III межд. конф. по лесному почвоведению. Петрозаводск: КарНЦ РАН, 2009. С. 199-203.
7. , А. Панцирные клещи (Acariformes, Oribatei) в таежных и тундровых почвах Мурманской области // Биогеография почв: Тез. II Всерос. конф. посвященной 70-летию со дня рождения . М.: МГУ, 2009. С. 34.
8. , А. Оценка эффективности способов восстановления техногенной пустоши в окрестностях медно-никелевого комбината «Североникель» по состоянию комплексов микроартропод // Современные проблемы загрязнения почв: Матер. III межд. науч. конф., М.: МГУ, 2010. С. 508-512.
9. , А. Почвообитающие беспозвоночные как индикаторы восстановления техногенной пустоши в окрестностях комбината «Североникель» (г. Мончегорск) // Сбалансированное природопользование: Матер. V Школы молодых ученых и специалистов КНЦ РАН. Апатиты: Изд-во КНЦ РАН, 2010. С. 41-47.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
